

Serine one-carbon catabolism with formate overflow
- PMID: 27819051
- PMCID: PMC5091358
- DOI: 10.1126/sciadv.1601273
Serine one-carbon catabolism with formate overflow
Abstract
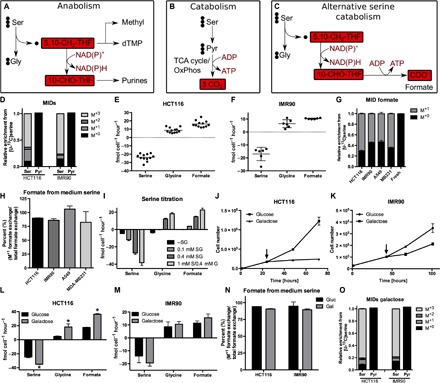
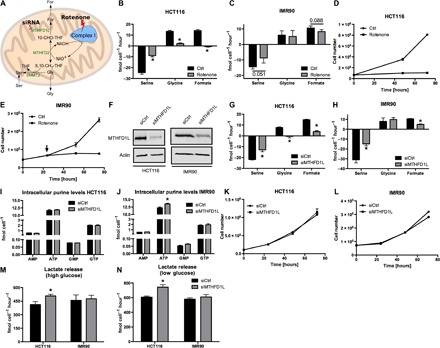
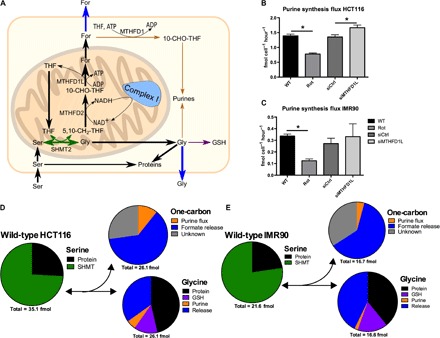
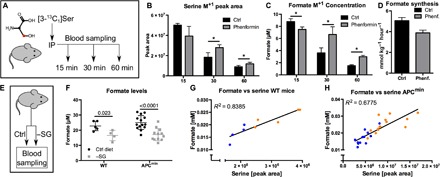
Serine catabolism to glycine and a one-carbon unit has been linked to the anabolic requirements of proliferating mammalian cells. However, genome-scale modeling predicts a catabolic role with one-carbon release as formate. We experimentally prove that in cultured cancer cells and nontransformed fibroblasts, most of the serine-derived one-carbon units are released from cells as formate, and that formate release is dependent on mitochondrial reverse 10-CHO-THF synthetase activity. We also show that in cancer cells, formate release is coupled to mitochondrial complex I activity, whereas in nontransformed fibroblasts, it is partially insensitive to inhibition of complex I activity. We demonstrate that in mice, about 50% of plasma formate is derived from serine and that serine starvation or complex I inhibition reduces formate synthesis in vivo. These observations transform our understanding of one-carbon metabolism and have implications for the treatment of diabetes and cancer with complex I inhibitors.
Keywords: Serine metabolism; folate metabolism; metformin; mitochondria metabolism; one-carbon metabolism; overflow metabolism.
Figures
References
-
- Tibbetts A. S., Appling D. R., Compartmentalization of mammalian folate-mediated one-carbon metabolism. Annu. Rev. Nutr. 30, 57–81 (2010). - PubMed
-
- Allen R. W., Moskowitz M., Arrest of cell growth in G1 phase of cell cycle by serine deprivation. Exp. Cell Res. 116, 127–137 (1978). - PubMed
-
- Rowe P. B., Sauer D., Fahey D., Craig G., Mccairns E., One-carbon metabolism in lectin-activated human-lymphocytes. Arch. Biochem. Biophys. 236, 277–288 (1985). - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
