

Saponin-based adjuvants induce cross-presentation in dendritic cells by intracellular lipid body formation
- PMID: 27819292
- PMCID: PMC5103066
- DOI: 10.1038/ncomms13324
Saponin-based adjuvants induce cross-presentation in dendritic cells by intracellular lipid body formation
Abstract
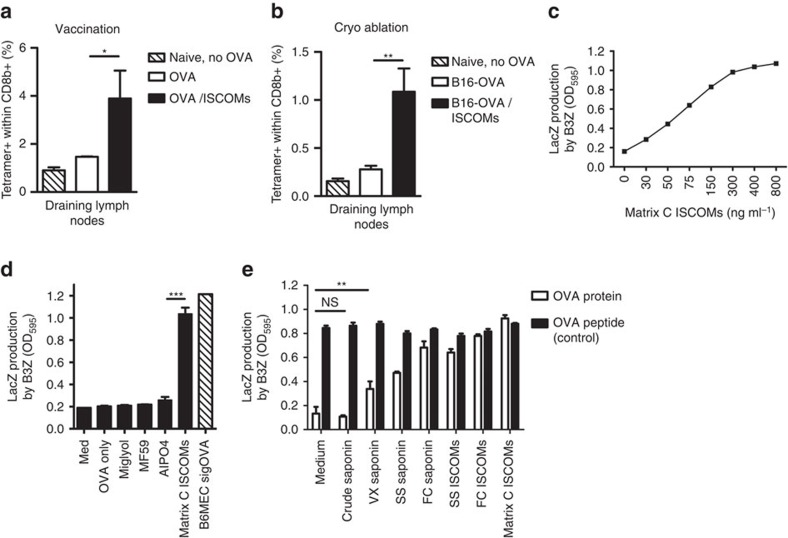
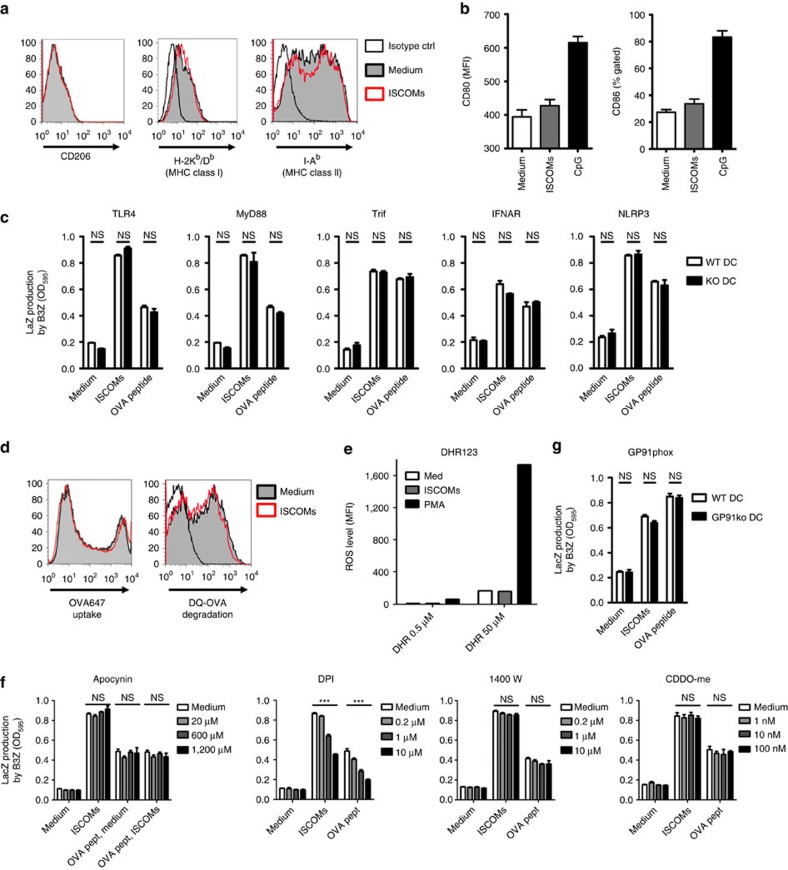
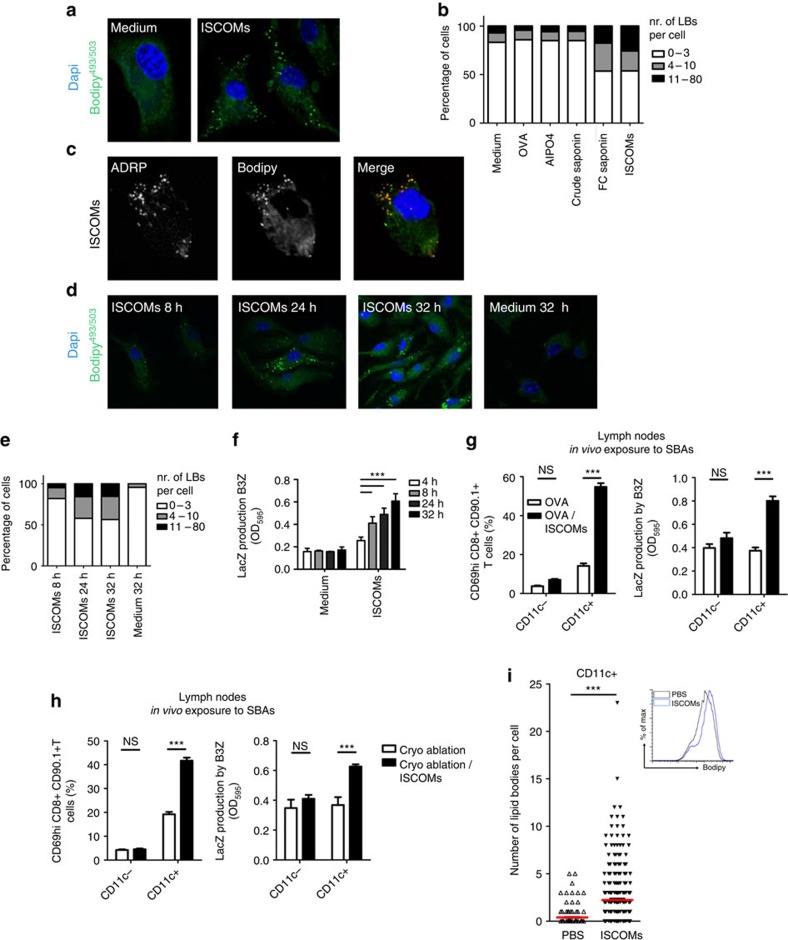
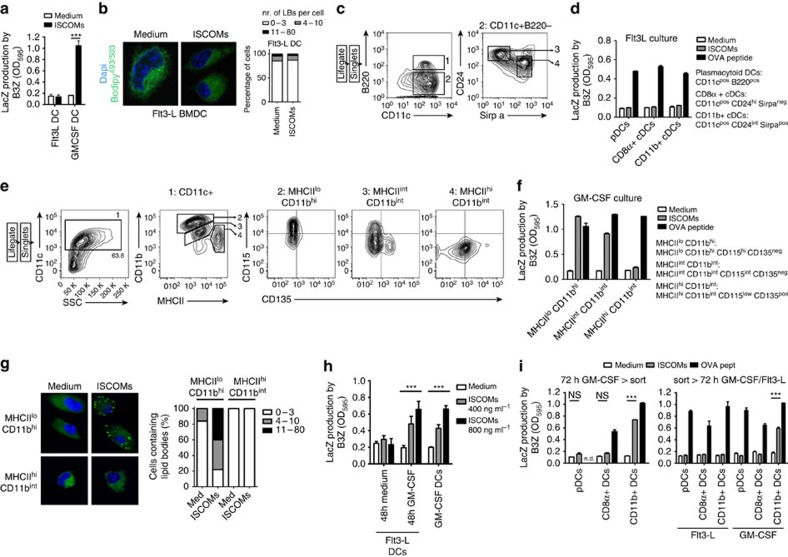
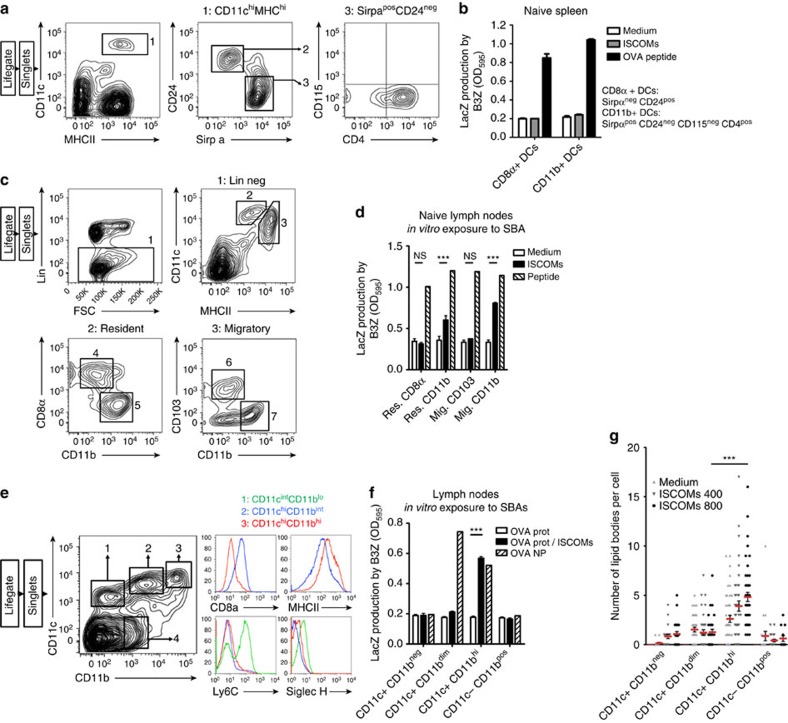
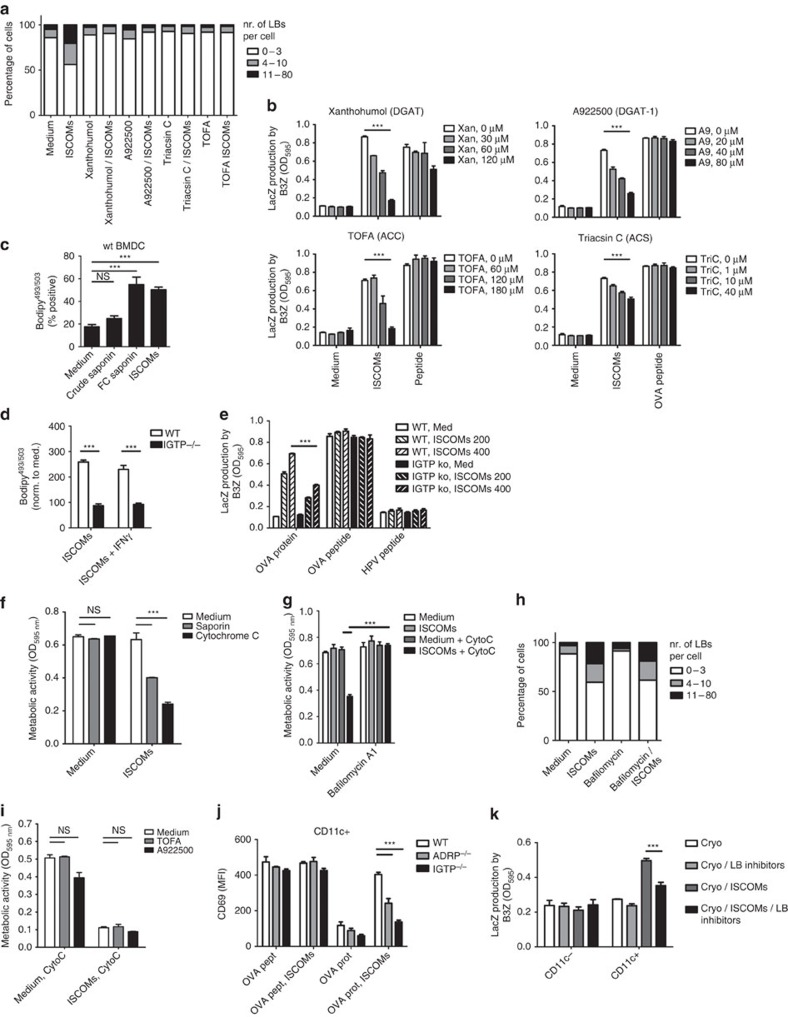
Saponin-based adjuvants (SBAs) are being used in animal and human (cancer) vaccines, as they induce protective cellular immunity. Their adjuvant potency is a factor of inflammasome activation and enhanced antigen cross-presentation by dendritic cells (DCs), but how antigen cross-presentation is induced is not clear. Here we show that SBAs uniquely induce intracellular lipid bodies (LBs) in the CD11b+ DC subset in vitro and in vivo. Using genetic and pharmacological interference in models for vaccination and in situ tumour ablation, we demonstrate that LB induction is causally related to the saponin-dependent increase in cross-presentation and T-cell activation. These findings link adjuvant activity to LB formation, aid the application of SBAs as a cancer vaccine component, and will stimulate development of new adjuvants enhancing T-cell-mediated immunity.
Figures
References
-
- Kensil C. R., Patel U., Lennick M. & Marciani D. Separation and characterization of saponins with adjuvant activity from Quillaja saponaria Molina cortex. J. Immunol. 146, 431–437 (1991). - PubMed
-
- Lorent J. H., Quetin-Leclercq J. & Mingeot-Leclercq M. P. The amphiphilic nature of saponins and their effects on artificial and biological membranes and potential consequences for red blood and cancer cells. Org. Biomol. Chem. 12, 8803–8822 (2014). - PubMed
-
- Morein B., Sundquist B., Hoglund S., Dalsgaard K. & Osterhaus A. Iscom, a novel structure for antigenic presentation of membrane proteins from enveloped viruses. Nature 308, 457–460 (1984). - PubMed
-
- Lovgren Bengtsson K., Morein B. & Osterhaus A. D. ISCOM technology-based Matrix M adjuvant: success in future vaccines relies on formulation. Expert Rev. Vaccines 10, 401–403 (2011). - PubMed
-
- Maraskovsky E. et al.. Development of prophylactic and therapeutic vaccines using the ISCOMATRIX adjuvant. Immunol. Cell Biol. 87, 371–376 (2009). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
