

Measurement of bacterial replication rates in microbial communities
- PMID: 27819664
- PMCID: PMC5538567
- DOI: 10.1038/nbt.3704
Measurement of bacterial replication rates in microbial communities
Abstract
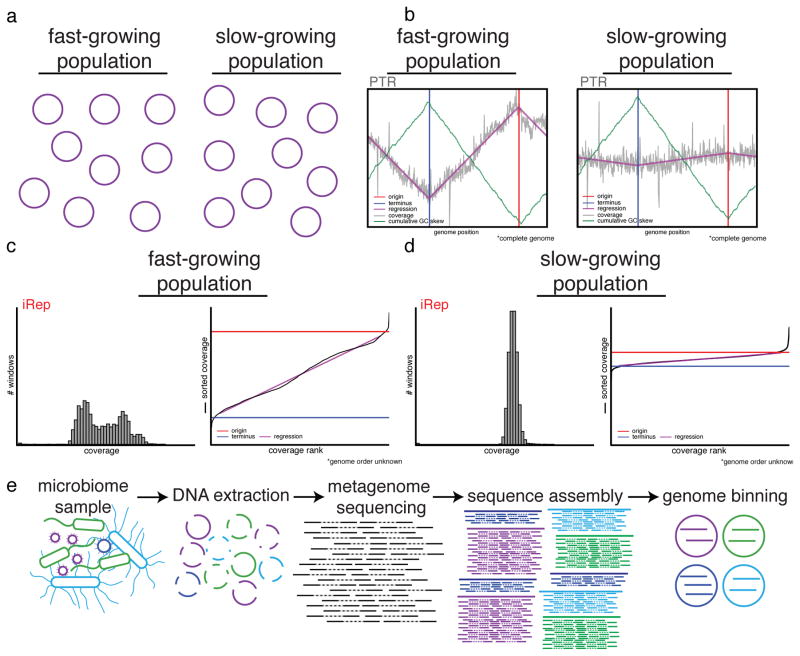
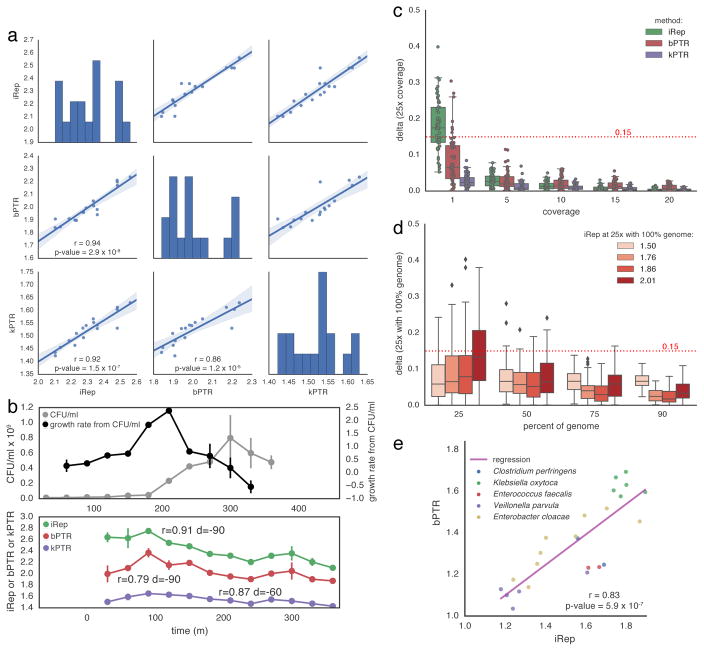
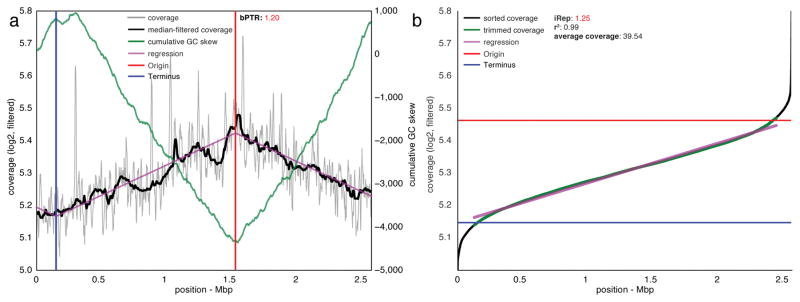
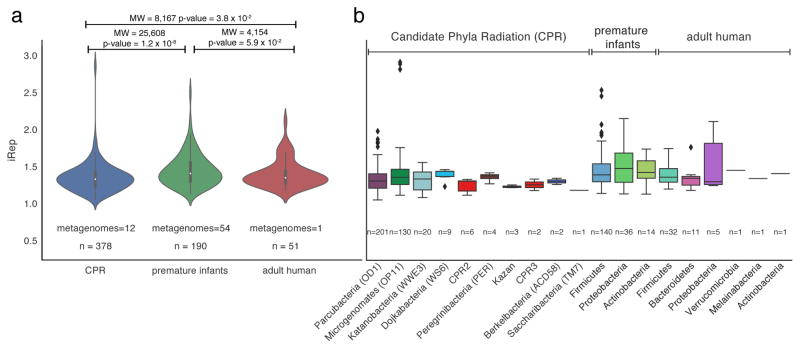
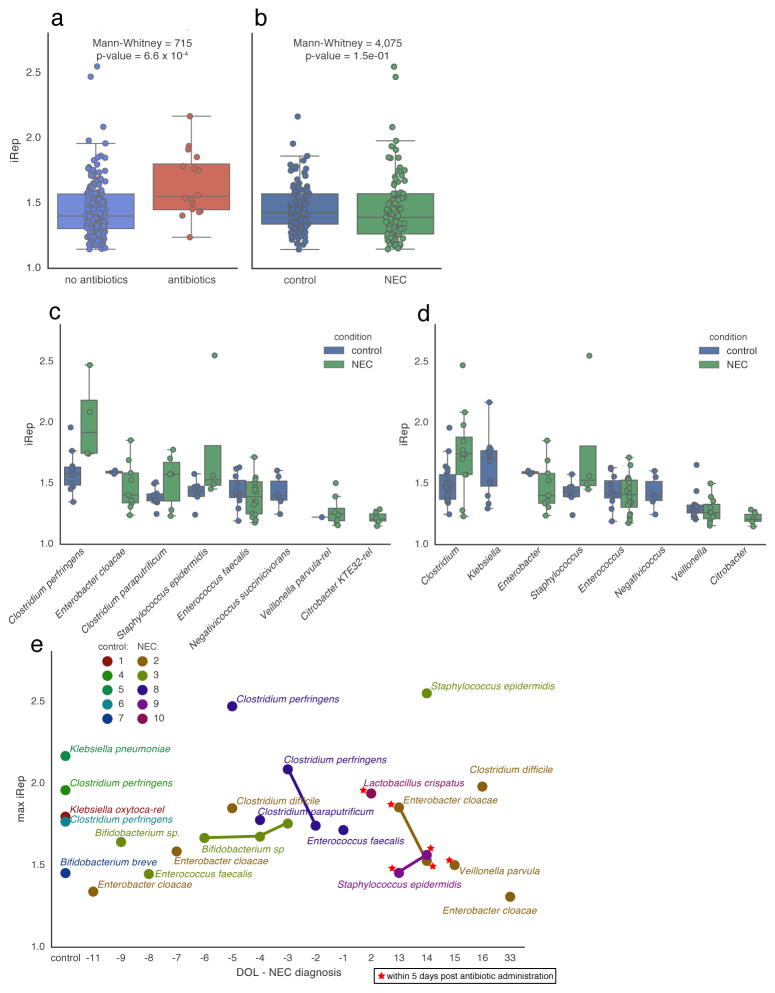
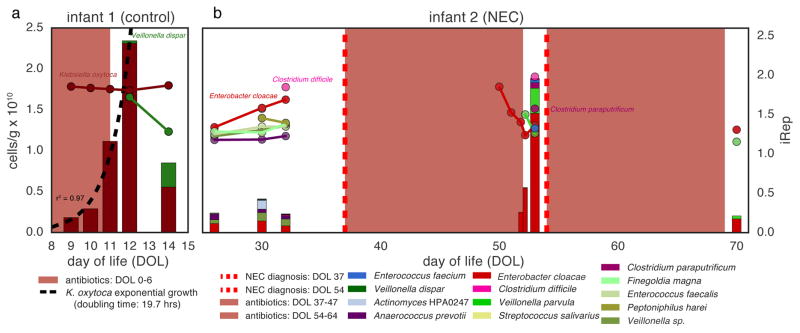
Culture-independent microbiome studies have increased our understanding of the complexity and metabolic potential of microbial communities. However, to understand the contribution of individual microbiome members to community functions, it is important to determine which bacteria are actively replicating. We developed an algorithm, iRep, that uses draft-quality genome sequences and single time-point metagenome sequencing to infer microbial population replication rates. The algorithm calculates an index of replication (iRep) based on the sequencing coverage trend that results from bi-directional genome replication from a single origin of replication. We apply this method to show that microbial replication rates increase after antibiotic administration in human infants. We also show that uncultivated, groundwater-associated, Candidate Phyla Radiation bacteria only rarely replicate quickly in subsurface communities undergoing substantial changes in geochemistry. Our method can be applied to any genome-resolved microbiome study to track organism responses to varying conditions, identify actively growing populations and measure replication rates for use in modeling studies.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Bremer H, Churchward G. An examination of the Cooper-Helmstetter theory of DNA replication in bacteria and its underlying assumptions. J Theor Biol. 1977;69:645–654. - PubMed
-
- Wake RG. Visualization of reinitiated chromosomes in Bacillus subtilis. J Mol Biol. 1972;68:501–509. - PubMed
-
- Sernova NV, Gelfand MS. Identification of replication origins in prokaryotic genomes. Briefings in Bioinformatics. 2008;9:376–391. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
