

Preclinical models for obesity research
- PMID: 27821603
- PMCID: PMC5117231
- DOI: 10.1242/dmm.026443
Preclinical models for obesity research
Abstract
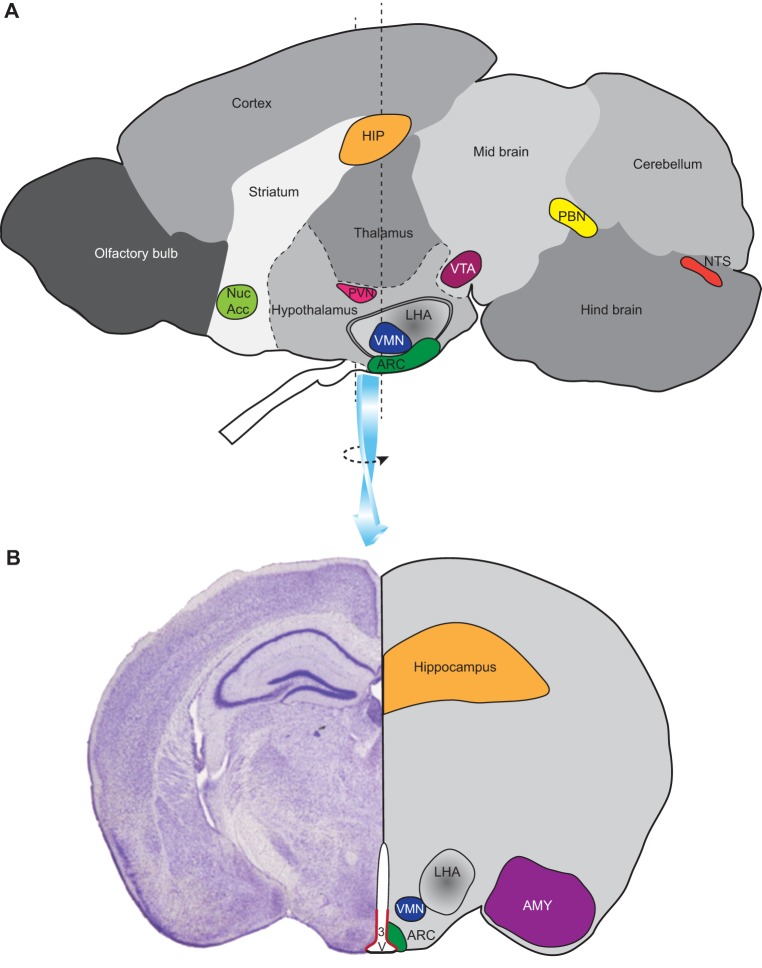
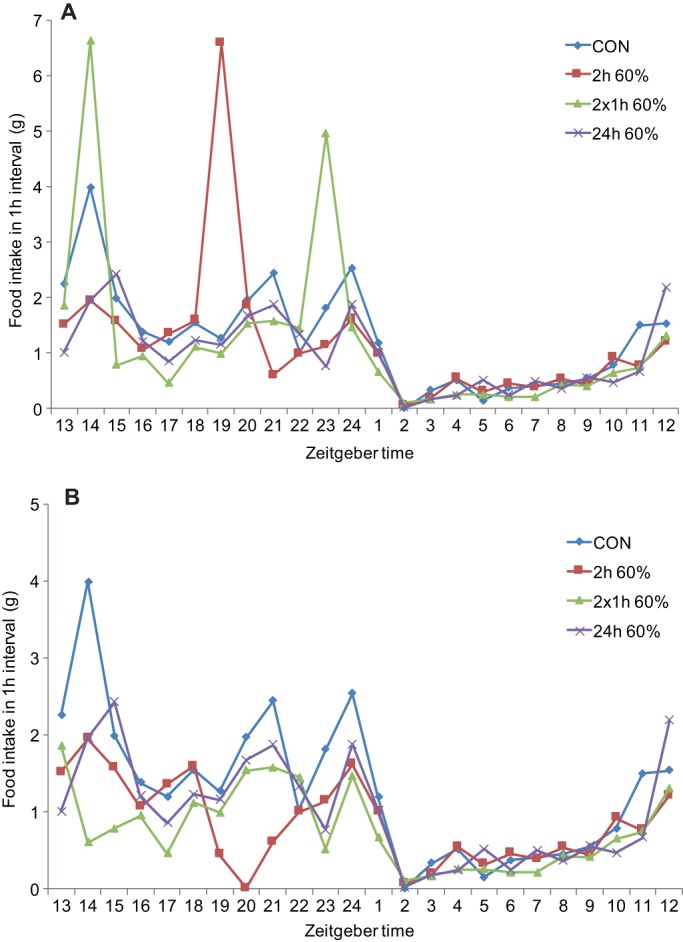
A multi-dimensional strategy to tackle the global obesity epidemic requires an in-depth understanding of the mechanisms that underlie this complex condition. Much of the current mechanistic knowledge has arisen from preclinical research performed mostly, but not exclusively, in laboratory mouse and rat strains. These experimental models mimic certain aspects of the human condition and its root causes, particularly the over-consumption of calories and unbalanced diets. As with human obesity, obesity in rodents is the result of complex gene-environment interactions. Here, we review the traditional monogenic models of obesity, their contemporary optogenetic and chemogenetic successors, and the use of dietary manipulations and meal-feeding regimes to recapitulate the complexity of human obesity. We critically appraise the strengths and weaknesses of these different models to explore the underlying mechanisms, including the neural circuits that drive behaviours such as appetite control. We also discuss the use of these models for testing and screening anti-obesity drugs, beneficial bio-actives, and nutritional strategies, with the goal of ultimately translating these findings for the treatment of human obesity.
Keywords: Binge eating; Epigenetics; Obesity; Transgenics.
© 2016. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
