

RSK2 activity mediates glioblastoma invasiveness and is a potential target for new therapeutics
- PMID: 27829215
- PMCID: PMC5346757
- DOI: 10.18632/oncotarget.13084
RSK2 activity mediates glioblastoma invasiveness and is a potential target for new therapeutics
Abstract
In glioblastoma (GBM), infiltration of primary tumor cells into the normal tissue and dispersal throughout the brain is a central challenge to successful treatment that remains unmet. Indeed, patients respond poorly to the current therapies of tumor resection followed by chemotherapy with radiotherapy and have only a 16-month median survival. It is therefore imperative to develop novel therapies. RSK2 is a kinase that regulates proliferation and adhesion and can promote metastasis. We demonstrate that active RSK2 regulates GBM cell adhesion and is essential for cell motility and invasion of patient-derived GBM neurospheres. RSK2 control of adhesion and migration is mediated in part by its effects on integrin-Filamin A complexes. Importantly, inhibition of RSK2 by either RSK inhibitors or shRNA silencing impairs invasion and combining RSK2 inhibitors with temozolomide improves efficacy in vitro. In agreement with the in vitro data, using public datasets, we find that RSK2 is significantly upregulated in vivo in human GBM patient tumors, and that high RSK2 expression significantly correlates with advanced tumor stage and poor patient survival. Together, our data provide strong evidence that RSK inhibitors could enhance the effectiveness of existing GBM treatment, and support RSK2 targeting as a promising approach for novel GBM therapy.
Keywords: ERK; GBM; MAPK; RSK; glioblastoma; invasion.
Conflict of interest statement
The authors declare that they have no competing financial interests.
Figures
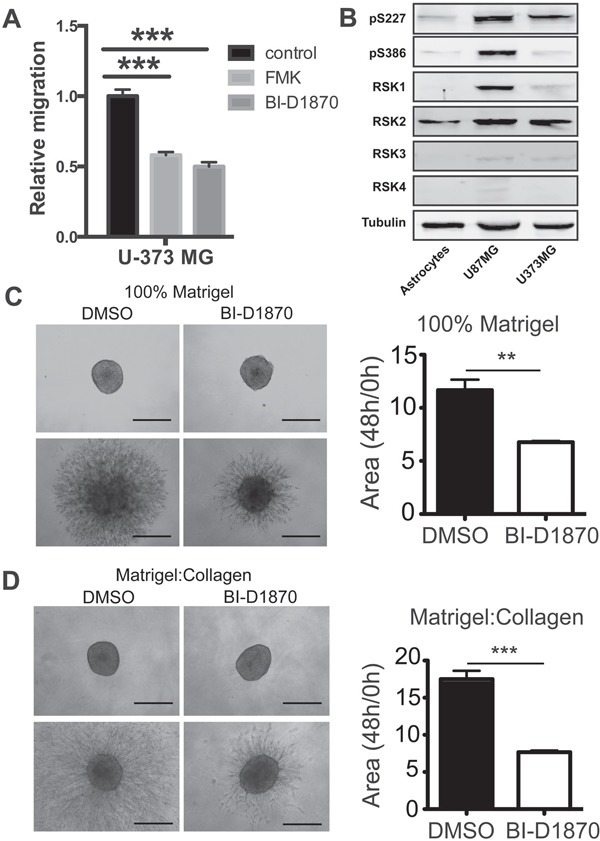
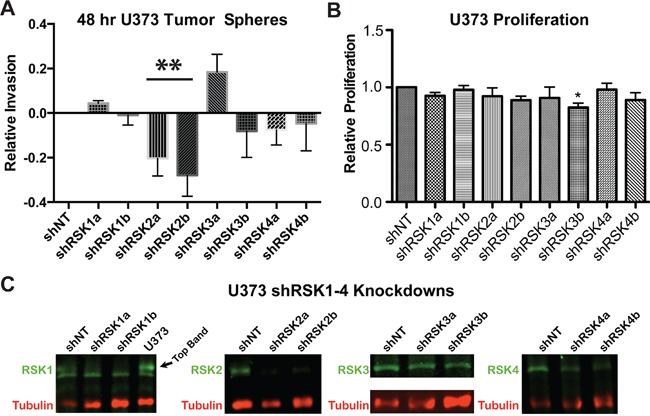
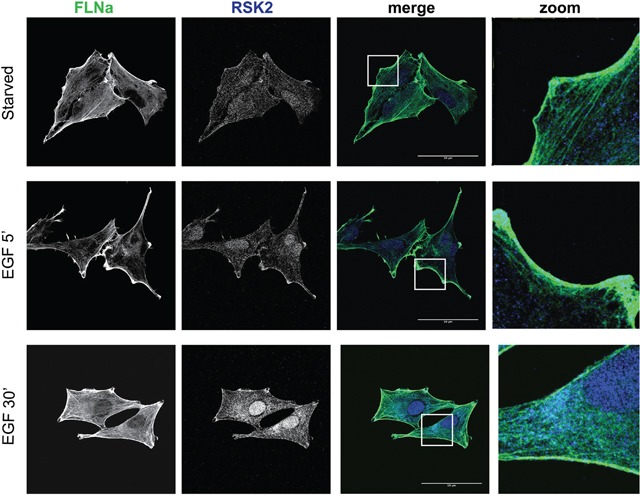
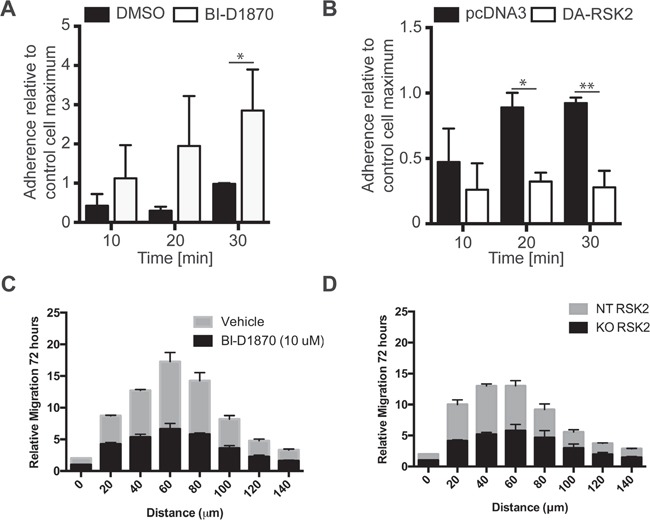
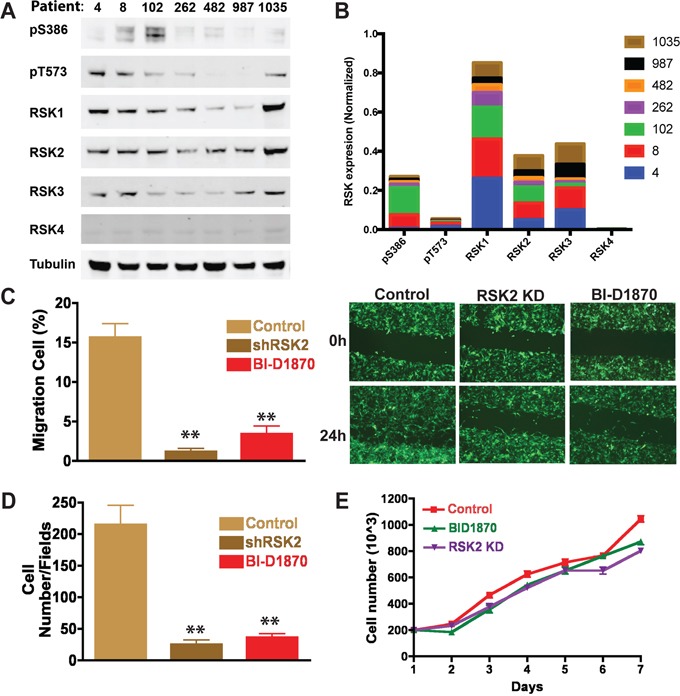
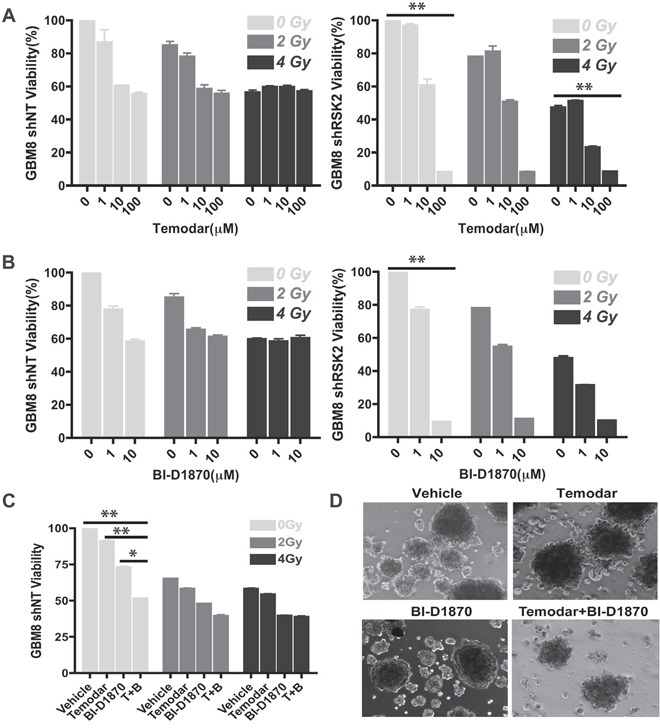
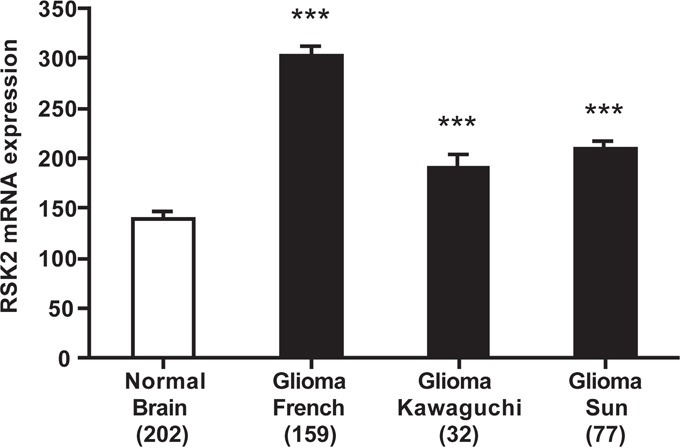
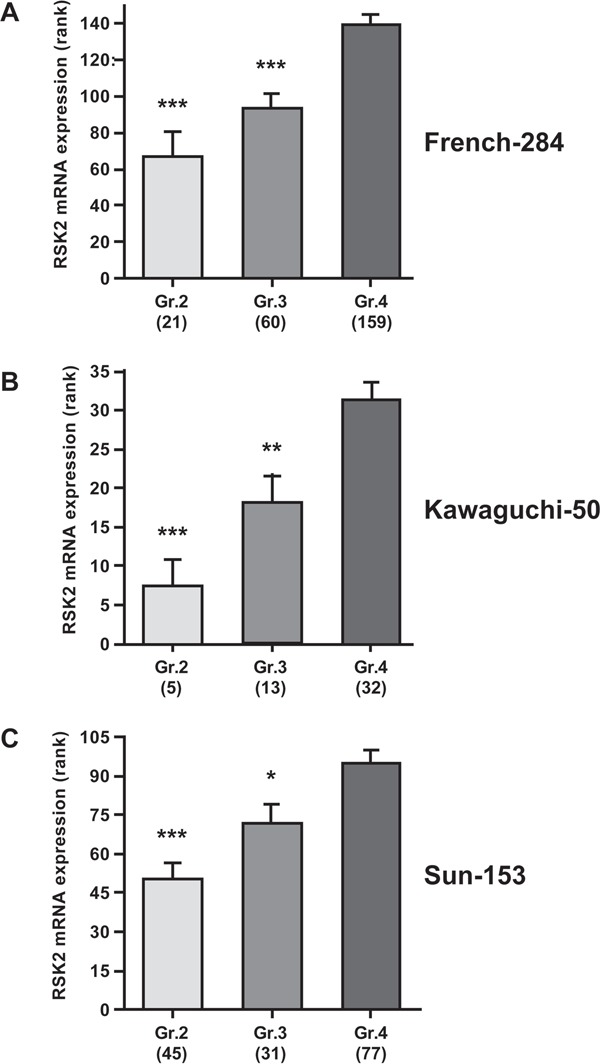

References
-
- Ignatova TN, Kukekov VG, Laywell ED, Suslov ON, Vrionis FD, Steindler DA. Human cortical glial tumors contain neural stem-like cells expressing astroglial and neuronal markers in vitro. Glia. 2002;39:193–206. - PubMed
-
- Wen PY, Kesari S. Malignant gliomas in adults. N Engl J Med. 2008;359:492–507. - PubMed
-
- Choucair AK, Levin VA, Gutin PH, Davis RL, Silver P, Edwards MS, Wilson CB. Development of multiple lesions during radiation therapy and chemotherapy in patients with gliomas. J Neurosurg. 1986;65:654–658. - PubMed
-
- Bartek J, Jr, Ng K, Bartek J, Fischer W, Carter B, Chen CC. Key concepts in glioblastoma therapy. J Neurol Neurosurg Psychiatry. 2012;83:753–760. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
