

Network abnormalities and interneuron dysfunction in Alzheimer disease
- PMID: 27829687
- PMCID: PMC8162106
- DOI: 10.1038/nrn.2016.141
Network abnormalities and interneuron dysfunction in Alzheimer disease
Abstract
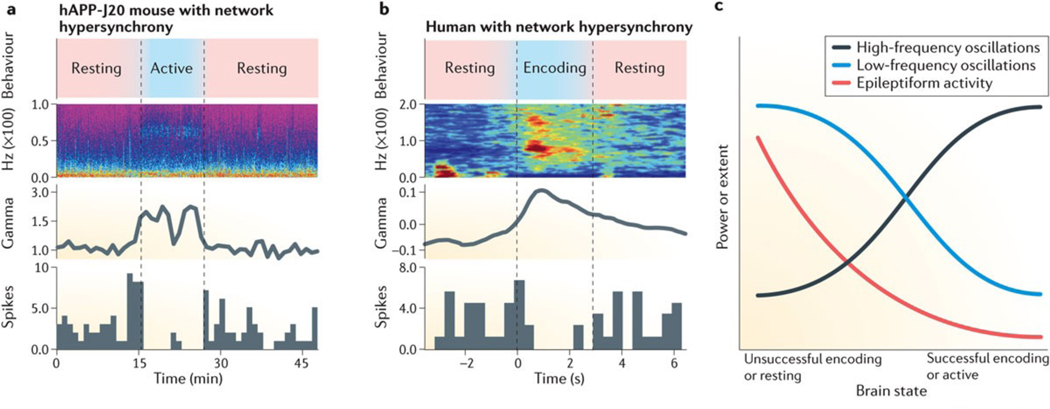
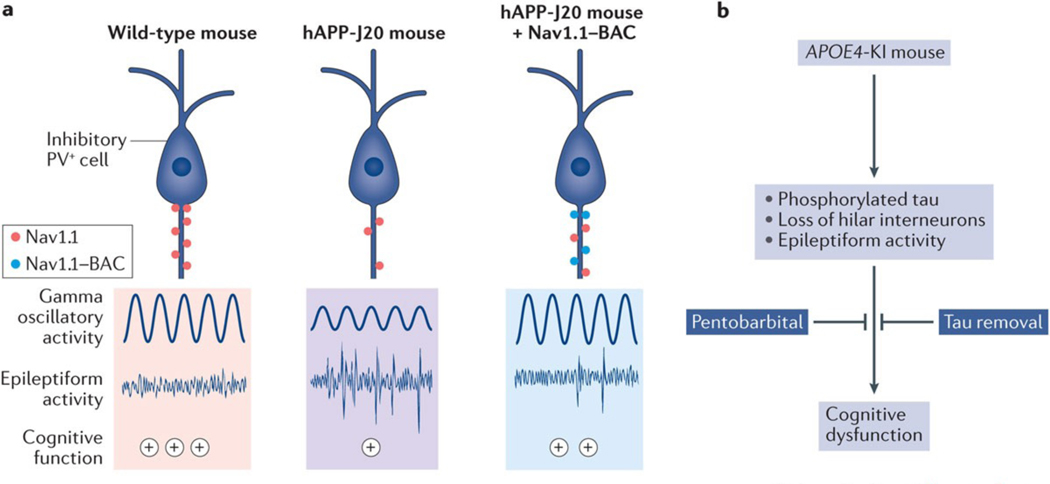
The function of neural circuits and networks can be controlled, in part, by modulating the synchrony of their components' activities. Network hypersynchrony and altered oscillatory rhythmic activity may contribute to cognitive abnormalities in Alzheimer disease (AD). In this condition, network activities that support cognition are altered decades before clinical disease onset, and these alterations predict future pathology and brain atrophy. Although the precise causes and pathophysiological consequences of these network alterations remain to be defined, interneuron dysfunction and network abnormalities have emerged as potential mechanisms of cognitive dysfunction in AD and related disorders. Here, we explore the concept that modulating these mechanisms may help to improve brain function in these conditions.
Figures
References
-
- Seidman LJ et al. Medial temporal lobe default mode functioning and hippocampal structure as vulnerability indicators for schizophrenia: A MRI study of non-psychotic adolescent first-degree relatives. Schizophr. Res 159, 426–434 (2014). - PubMed
-
- Oser N. et al. Default mode network alterations during language task performance in children with benign epilepsy with centrotemporal spikes (BECTS). Epilepsy Behav. 33, 12–17 (2014). - PubMed
-
-
Sperling RA et al. Amyloid deposition is associated with impaired default network function in older persons without dementia. Neuron 63, 178–188 (2009).
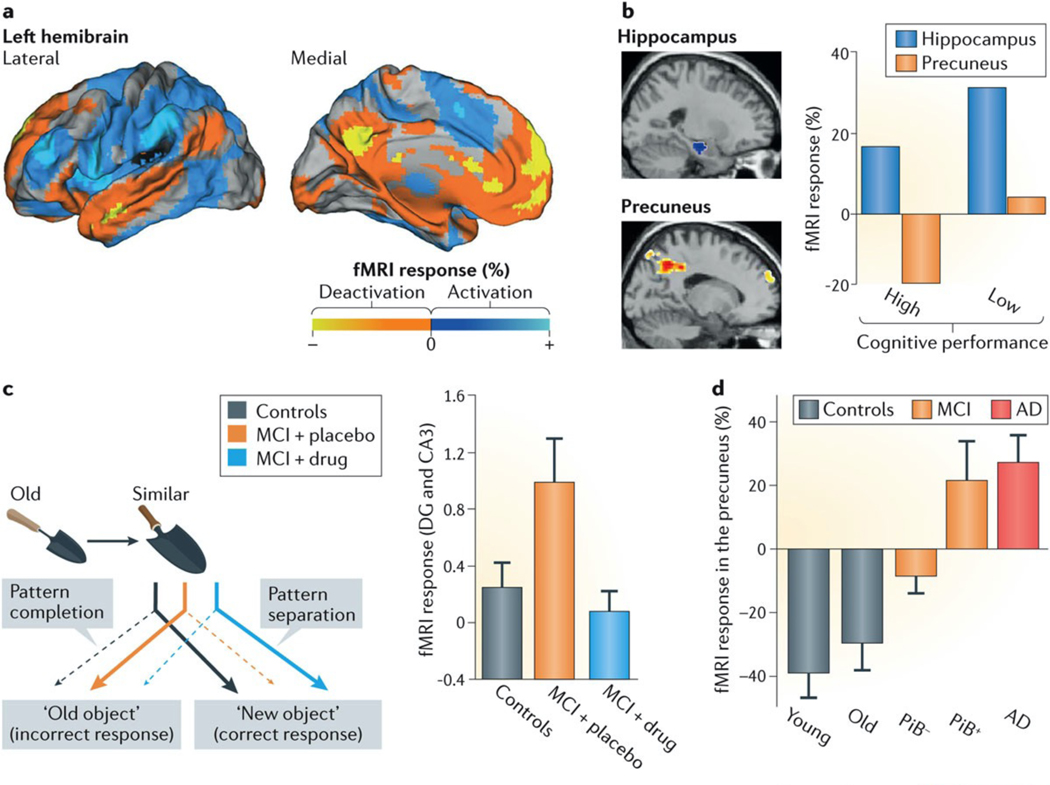
This study shows that PiB+ amyloid deposits are associated with deactivation deficits in a default network component in non-demented individuals and patients with MCI, linking aberrant neuronal activity and pathological changes.
-
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
