

Transcriptional start site heterogeneity modulates the structure and function of the HIV-1 genome
- PMID: 27834211
- PMCID: PMC5127319
- DOI: 10.1073/pnas.1616627113
Transcriptional start site heterogeneity modulates the structure and function of the HIV-1 genome
Abstract
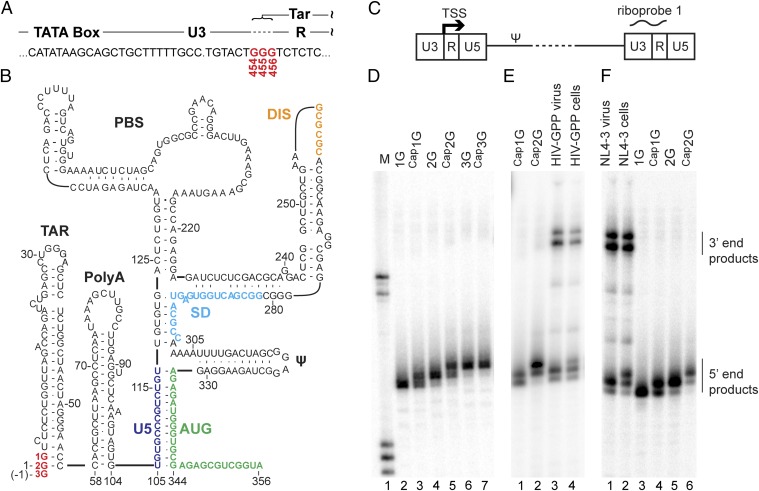
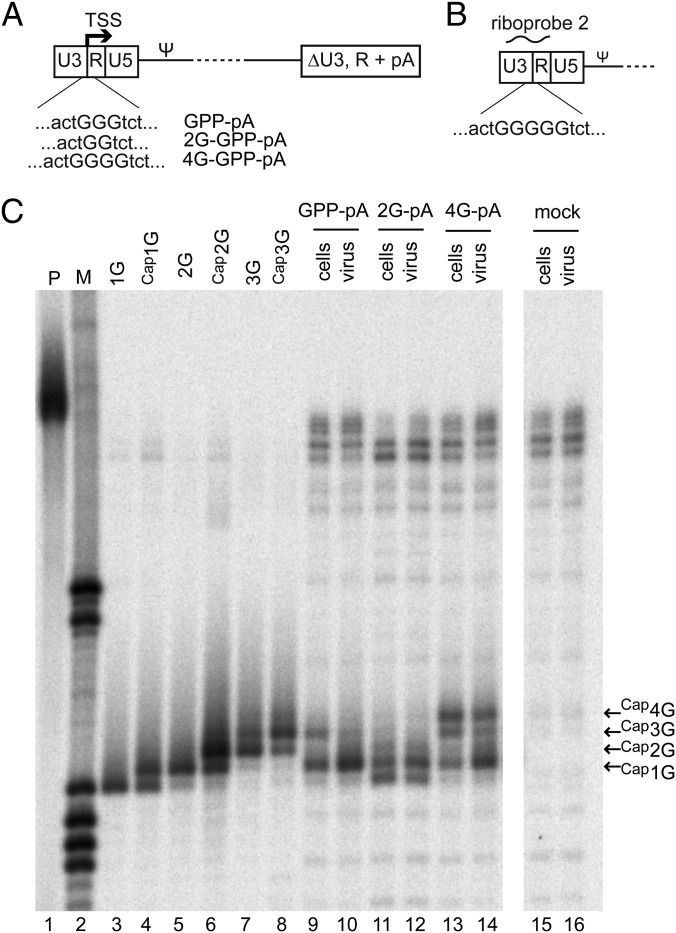
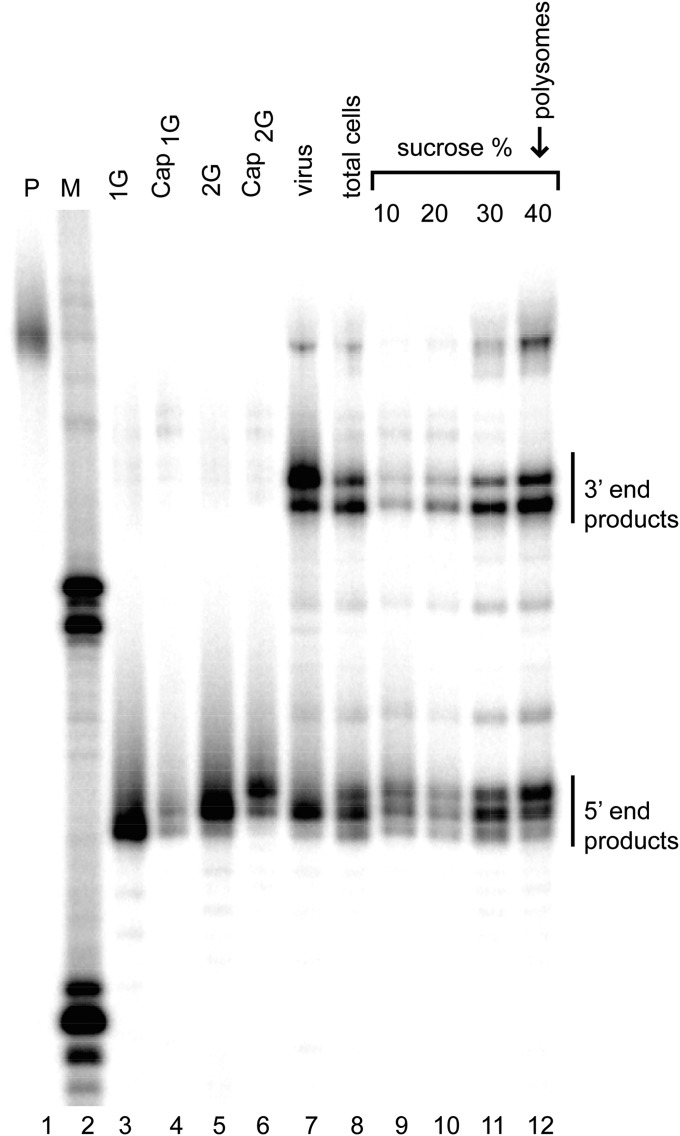
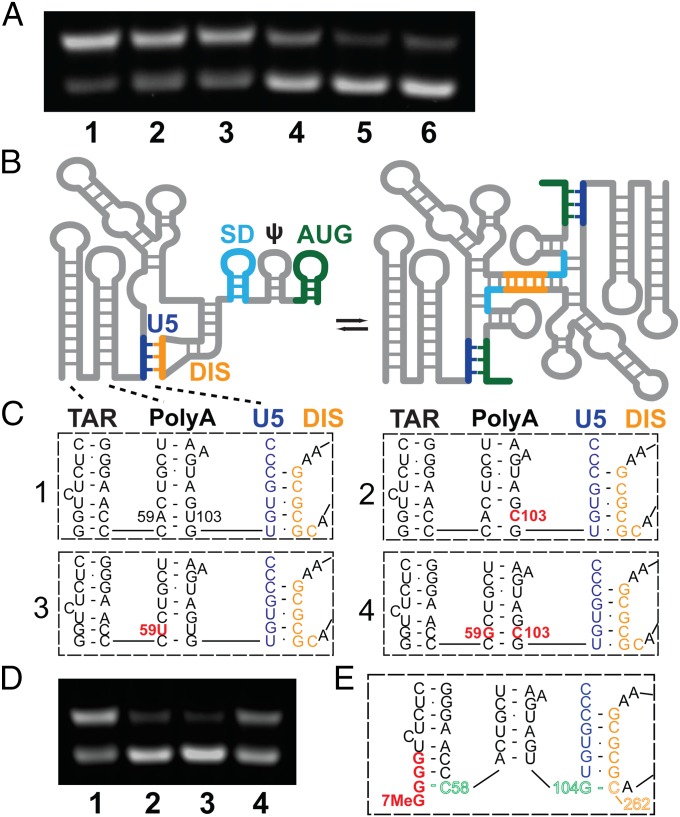
The promoter in HIV type 1 (HIV-1) proviral DNA contains three sequential guanosines at the U3-R boundary that have been proposed to function as sites for transcription initiation. Here we show that all three sites are used in cells infected with HIV-1 and that viral RNAs containing a single 5' capped guanosine (Cap1G) are specifically selected for packaging in virions, consistent with a recent report [Masuda et al. (2015) Sci Rep 5:17680]. In addition, we now show that transcripts that begin with two or three capped guanosines (Cap2G or Cap3G) are enriched on polysomes, indicating that RNAs synthesized from different transcription start sites have different functions in viral replication. Because genomes are selected for packaging as dimers, we examined the in vitro monomer-dimer equilibrium properties of Cap1G, Cap2G, and Cap3G 5'-leader RNAs in the NL4-3 strain of HIV-1. Strikingly, under physiological-like ionic conditions in which the Cap1G 5'-leader RNA adopts a dimeric structure, the Cap2G and Cap3G 5'-leader RNAs exist predominantly as monomers. Mutagenesis studies designed to probe for base-pairing interactions suggest that the additional guanosines of the 2G and 3G RNAs remodel the base of the PolyA hairpin, resulting in enhanced sequestration of dimer-promoting residues and stabilization of the monomer. Our studies suggest a mechanism through which the structure, function, and fate of the viral genome can be modulated by the transcriptionally controlled presence or absence of a single 5' guanosine.
Keywords: 5′-leader; HIV-1; RNA; structure; transcription.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Coffin JM, Hughes SH, Varmus HE. Retroviruses. Cold Spring Harbor Lab Press; Plainview, NY: 1997. - PubMed
-
- Chiu YL, Coronel E, Ho CK, Shuman S, Rana TM. HIV-1 Tat protein interacts with mammalian capping enzyme and stimulates capping of TAR RNA. J Biol Chem. 2001;276(16):12959–12966. - PubMed
-
- Menees TM, Müller B, Kräusslich HG. The major 5′ end of HIV type 1 RNA corresponds to G456. AIDS Res Hum Retroviruses. 2007;23(8):1042–1048. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
