

Phosphorylated Groucho delays differentiation in the follicle stem cell lineage by providing a molecular memory of EGFR signaling in the niche
- PMID: 27836963
- PMCID: PMC5201033
- DOI: 10.1242/dev.143263
Phosphorylated Groucho delays differentiation in the follicle stem cell lineage by providing a molecular memory of EGFR signaling in the niche
Abstract
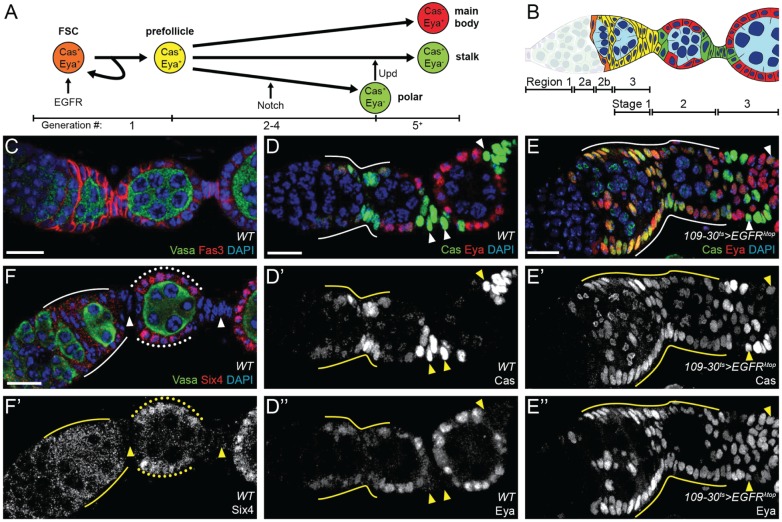
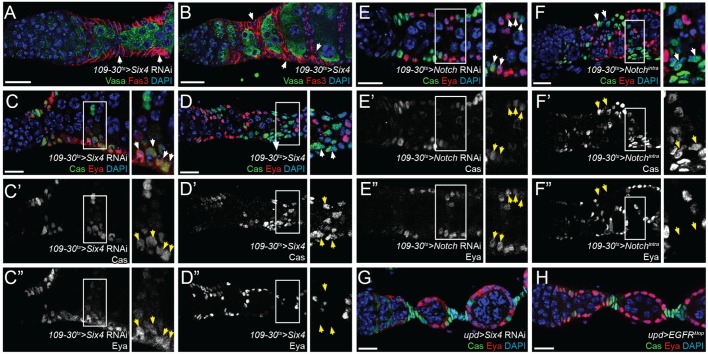
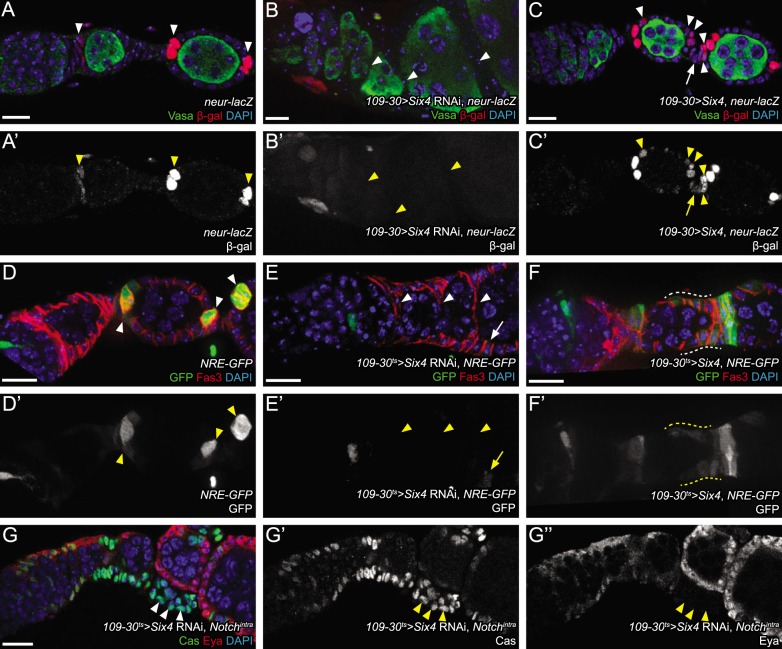
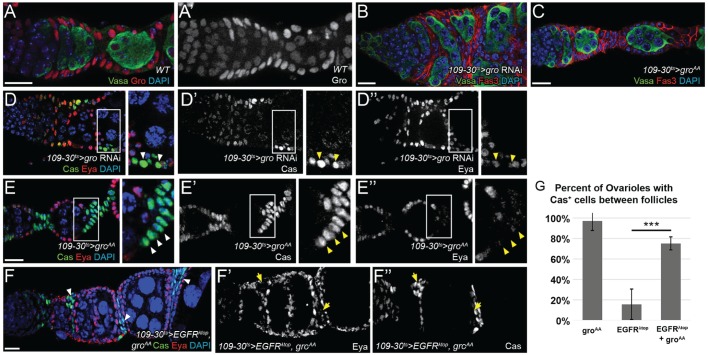
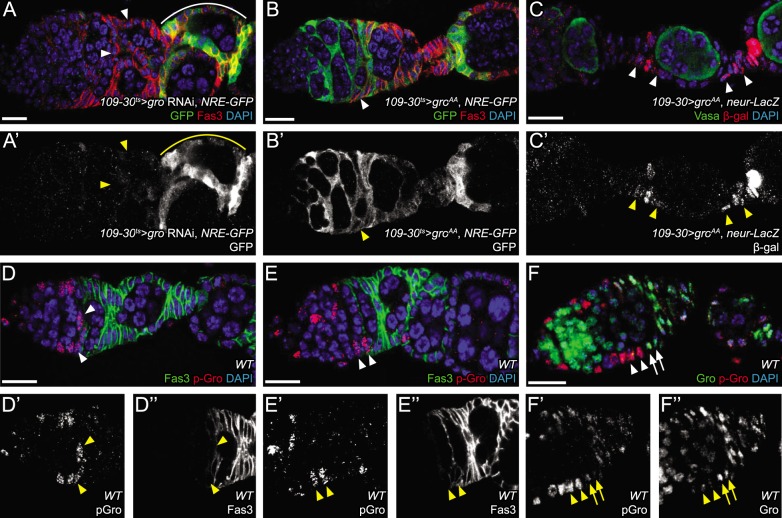
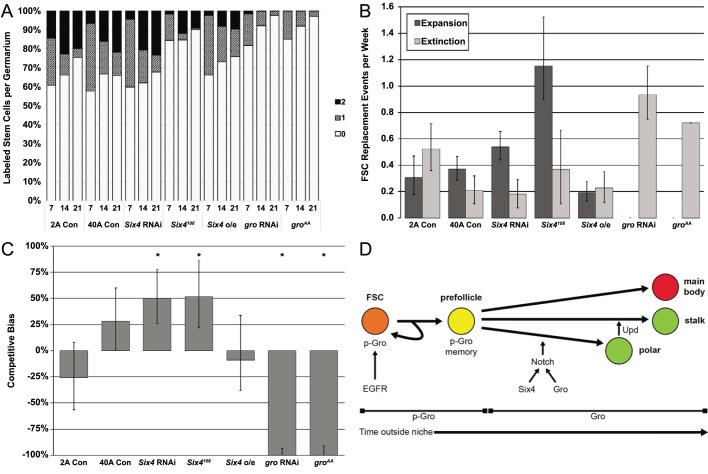
In the epithelial follicle stem cells (FSCs) of the Drosophila ovary, Epidermal Growth Factor Receptor (EGFR) signaling promotes self-renewal, whereas Notch signaling promotes differentiation of the prefollicle cell (pFC) daughters. We have identified two proteins, Six4 and Groucho (Gro), that link the activity of these two pathways to regulate the earliest cell fate decision in the FSC lineage. Our data indicate that Six4 and Gro promote differentiation towards the polar cell fate by promoting Notch pathway activity. This activity of Gro is antagonized by EGFR signaling, which inhibits Gro-dependent repression via p-ERK mediated phosphorylation. We have found that the phosphorylated form of Gro persists in newly formed pFCs, which may delay differentiation and provide these cells with a temporary memory of the EGFR signal. Collectively, these findings demonstrate that phosphorylated Gro labels a transition state in the FSC lineage and describe the interplay between Notch and EGFR signaling that governs the differentiation processes during this period.
Keywords: Drosophila; EGFR; Epithelial stem cell; Groucho; Ovary; Six4.
© 2016. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
Similar articles
-
Wingless promotes EGFR signaling in follicle stem cells to maintain self-renewal.Development. 2018 Dec 5;145(23):dev168716. doi: 10.1242/dev.168716. Development. 2018. PMID: 30389852 Free PMC article.
-
Groucho restricts rhomboid expression and couples EGFR activation with R8 selection during Drosophila photoreceptor differentiation.Dev Biol. 2015 Nov 15;407(2):246-55. doi: 10.1016/j.ydbio.2015.09.011. Epub 2015 Sep 28. Dev Biol. 2015. PMID: 26417727 Free PMC article.
-
Division of Labor: Roles of Groucho and CtBP in Notch-Mediated Lateral Inhibition that Controls Intestinal Stem Cell Differentiation in Drosophila.Stem Cell Reports. 2019 May 14;12(5):1007-1023. doi: 10.1016/j.stemcr.2019.03.005. Epub 2019 Apr 11. Stem Cell Reports. 2019. PMID: 30982741 Free PMC article.
-
Crosstalk between the EGFR and other signalling pathways at the level of the global transcriptional corepressor Groucho/TLE.Br J Cancer. 2006 Mar 27;94(6):771-5. doi: 10.1038/sj.bjc.6603019. Br J Cancer. 2006. PMID: 16508633 Free PMC article. Review.
-
[Main components of gene network controlling development of dorsal appendages of egg chorion in Drosophila melanogaster].Ontogenez. 2012 May-Jun;43(3):163-74. Ontogenez. 2012. PMID: 22834131 Review. Russian.
Cited by
-
Opposing JAK-STAT and Wnt signaling gradients define a stem cell domain by regulating differentiation at two borders.Elife. 2020 Nov 2;9:e61204. doi: 10.7554/eLife.61204. Elife. 2020. PMID: 33135631 Free PMC article.
-
Homeodomain protein Six4 prevents the generation of supernumerary Drosophila type II neuroblasts and premature differentiation of intermediate neural progenitors.PLoS Genet. 2021 Feb 8;17(2):e1009371. doi: 10.1371/journal.pgen.1009371. eCollection 2021 Feb. PLoS Genet. 2021. PMID: 33556050 Free PMC article.
-
IGF2BP3‑stabilized SIX4 promotes the proliferation, migration, invasion and tube formation of ovarian cancer cells.Mol Med Rep. 2022 Jul;26(1):232. doi: 10.3892/mmr.2022.12748. Epub 2022 May 26. Mol Med Rep. 2022. PMID: 35616130 Free PMC article.
-
Normal cell cycle progression requires negative regulation of E2F1 by Groucho during S phase and its relief at G2 phase.Development. 2023 Jun 1;150(11):dev201041. doi: 10.1242/dev.201041. Epub 2023 Jun 1. Development. 2023. PMID: 37260146 Free PMC article.
-
Drosophila Glypicans Regulate Follicle Stem Cell Maintenance and Niche Competition.Genetics. 2018 Jun;209(2):537-549. doi: 10.1534/genetics.118.300839. Epub 2018 Apr 9. Genetics. 2018. PMID: 29632032 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
