

Mitogen-Activated Protein Kinase Phosphatases Affect UV-B-Induced Stomatal Closure via Controlling NO in Guard Cells
- PMID: 27837091
- PMCID: PMC5210765
- DOI: 10.1104/pp.16.01656
Mitogen-Activated Protein Kinase Phosphatases Affect UV-B-Induced Stomatal Closure via Controlling NO in Guard Cells
Abstract
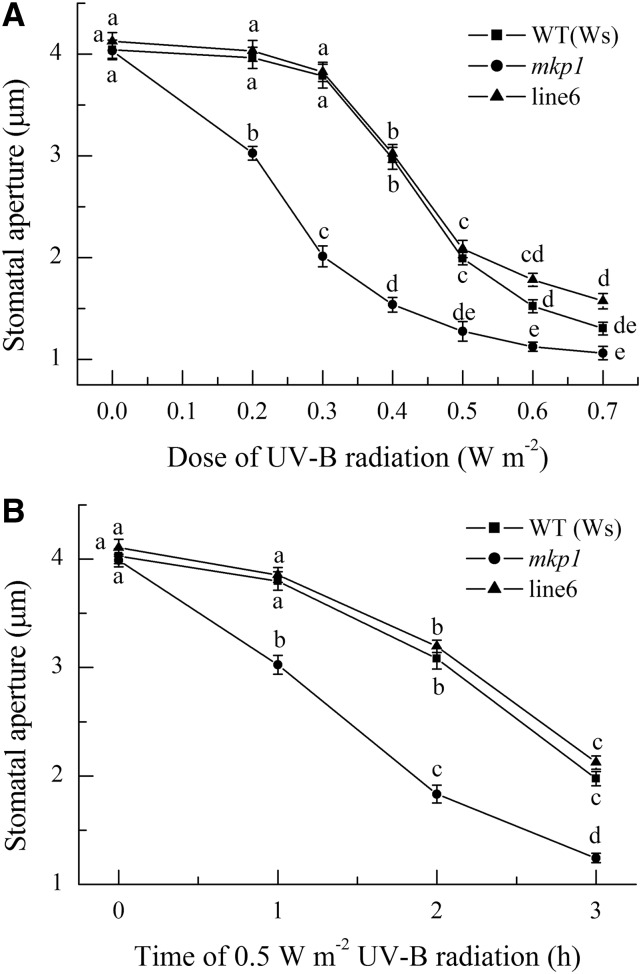
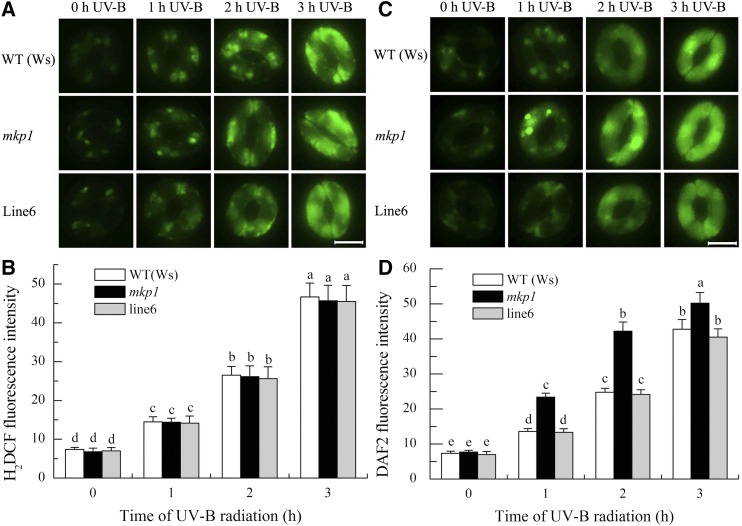
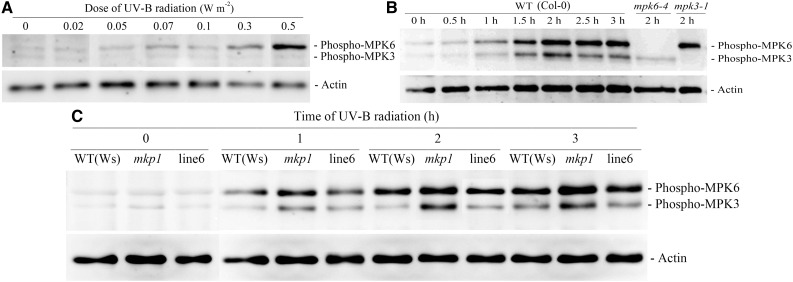
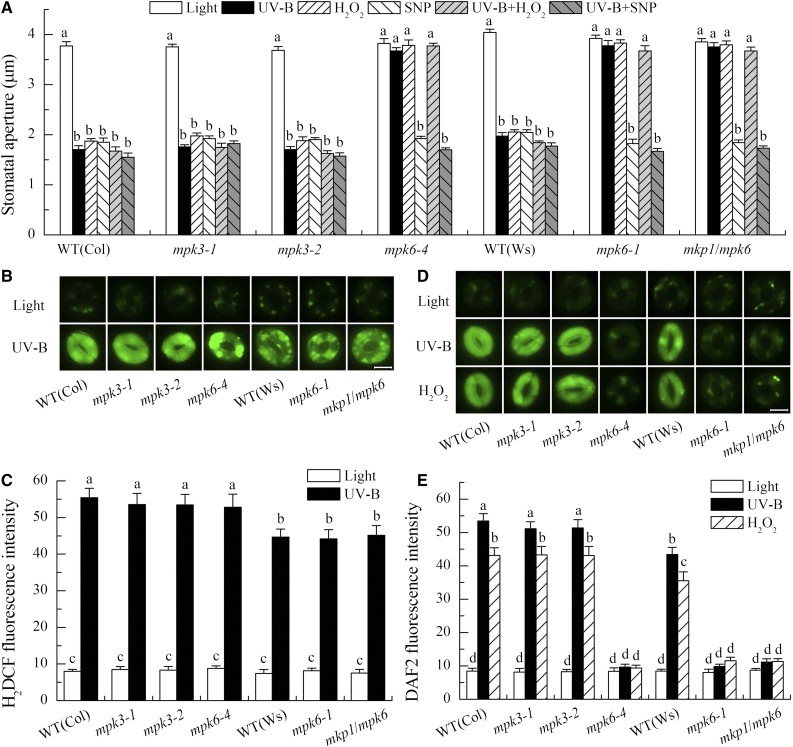
Ultraviolet B (UV-B) radiation induces the activation of MITOGEN-ACTIVATED PROTEIN KINASE PHOSPHATASE1 (MKP1) and its targets MPK3 and MPK6, but whether they participate in UV-B guard cell signaling is not clear. Here, evidence shows that UV-B-induced stomatal closure in Arabidopsis (Arabidopsis thaliana) is antagonistically regulated by MKP1 and MPK6 via modulating hydrogen peroxide (H2O2)-induced nitric oxide (NO) production in guard cells. The mkp1 mutant was hypersensitive to UV-B-induced stomatal closure and NO production in guard cells but not to UV-B-induced H2O2 production, suggesting that MKP1 negatively regulates UV-B-induced stomatal closure via inhibiting NO generation in guard cells. Moreover, MPK3 and MPK6 were activated by UV-B in leaves of the wild type and hyperactivated in the mkp1 mutant, but the UV-B-induced activation of MPK3 and MPK6 was largely inhibited in mutants for ATRBOHD and ATRBOHF but not in mutants for NIA1 and NIA2 mpk6 mutants showed defects of UV-B-induced NO production and stomatal closure but were normal in UV-B-induced H2O2 production, while stomata of mpk3 mutants responded to UV-B just like those of the wild type. The defect of UV-B-induced stomatal closure in mpk6 mutants was rescued by exogenous NO but not by exogenous H2O2 Furthermore, double mutant mkp1/mpk6 and the single mutant mpk6 showed the same responses to UV-B in terms of either stomatal movement or H2O2 and NO production. These data indicate that MPK6, but not MPK3, positively regulates UV-B-induced stomatal closure via acting downstream of H2O2 and upstream of NO, while MKP1 functions negatively in UV-B guard cell signaling via down-regulation of MPK6.
© 2017 American Society of Plant Biologists. All Rights Reserved.
Figures

Similar articles
-
MAP KINASE PHOSPHATASE1 Controls Cell Fate Transition during Stomatal Development.Plant Physiol. 2018 Sep;178(1):247-257. doi: 10.1104/pp.18.00475. Epub 2018 Jul 12. Plant Physiol. 2018. PMID: 30002258 Free PMC article.
-
Role and interrelationship of MEK1-MPK6 cascade, hydrogen peroxide and nitric oxide in darkness-induced stomatal closure.Plant Sci. 2017 Sep;262:190-199. doi: 10.1016/j.plantsci.2017.06.010. Epub 2017 Jul 1. Plant Sci. 2017. PMID: 28716416
-
Role and interrelationship of Gα protein, hydrogen peroxide, and nitric oxide in ultraviolet B-induced stomatal closure in Arabidopsis leaves.Plant Physiol. 2013 Mar;161(3):1570-83. doi: 10.1104/pp.112.211623. Epub 2013 Jan 22. Plant Physiol. 2013. PMID: 23341360 Free PMC article.
-
Nitric oxide, stomatal closure, and abiotic stress.J Exp Bot. 2008;59(2):165-76. doi: 10.1093/jxb/erm293. J Exp Bot. 2008. PMID: 18332225 Review.
-
Reactive Oxygen Species in the Regulation of Stomatal Movements.Plant Physiol. 2016 Jul;171(3):1569-80. doi: 10.1104/pp.16.00328. Epub 2016 Apr 21. Plant Physiol. 2016. PMID: 27208297 Free PMC article. Review.
Cited by
-
Local endocytosis of sucrose transporter 2 in duckweed reveals the role of sucrose transporter 2 in guard cells.Front Plant Sci. 2022 Oct 24;13:996618. doi: 10.3389/fpls.2022.996618. eCollection 2022. Front Plant Sci. 2022. PMID: 36352881 Free PMC article.
-
MAP KINASE PHOSPHATASE1 Controls Cell Fate Transition during Stomatal Development.Plant Physiol. 2018 Sep;178(1):247-257. doi: 10.1104/pp.18.00475. Epub 2018 Jul 12. Plant Physiol. 2018. PMID: 30002258 Free PMC article.
-
The role of calcium-dependent protein kinase in hydrogen peroxide, nitric oxide and ABA-dependent cold acclimation.J Exp Bot. 2018 Jul 18;69(16):4127-4139. doi: 10.1093/jxb/ery212. J Exp Bot. 2018. PMID: 29868714 Free PMC article.
-
Free Radicals Mediated Redox Signaling in Plant Stress Tolerance.Life (Basel). 2023 Jan 10;13(1):204. doi: 10.3390/life13010204. Life (Basel). 2023. PMID: 36676153 Free PMC article. Review.
-
CRISPR/Cas9-mediated knockout of PiSSK1 reveals essential role of S-locus F-box protein-containing SCF complexes in recognition of non-self S-RNases during cross-compatible pollination in self-incompatible Petunia inflata.Plant Reprod. 2018 Jun;31(2):129-143. doi: 10.1007/s00497-017-0314-1. Epub 2017 Nov 30. Plant Reprod. 2018. PMID: 29192328
References
-
- Anderson JC, Bartels S, González Besteiro MA, Shahollari B, Ulm R, Peck SC (2011) Arabidopsis MAP Kinase Phosphatase 1 (AtMKP1) negatively regulates MPK6-mediated PAMP responses and resistance against bacteria. Plant J 67: 258–268 - PubMed
-
- Asai T, Tena G, Plotnikova J, Willmann MR, Chiu WL, Gomez-Gomez L, Boller T, Ausubel FM, Sheen J (2002) MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 415: 977–983 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
