

Odour-induced analgesia mediated by hypothalamic orexin neurons in mice
- PMID: 27845440
- PMCID: PMC5109046
- DOI: 10.1038/srep37129
Odour-induced analgesia mediated by hypothalamic orexin neurons in mice
Abstract
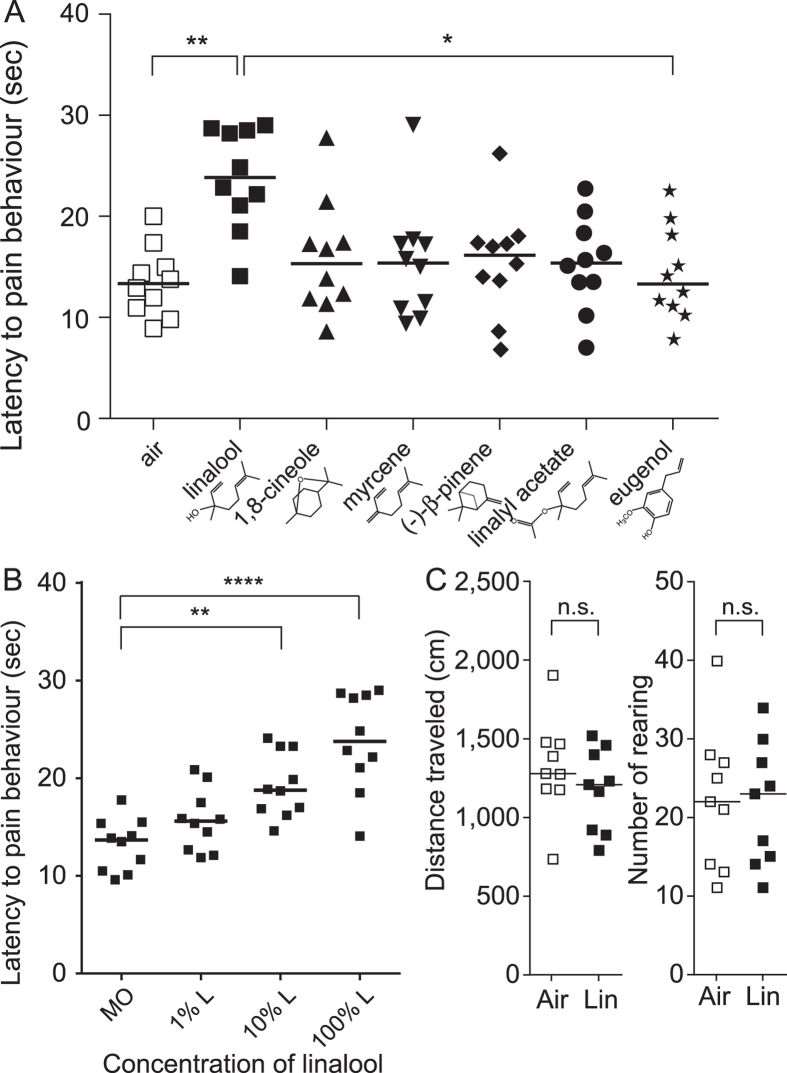
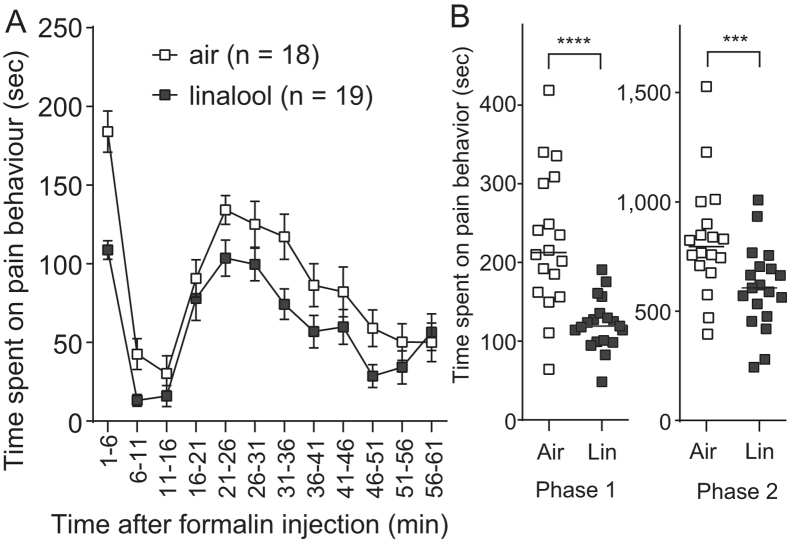
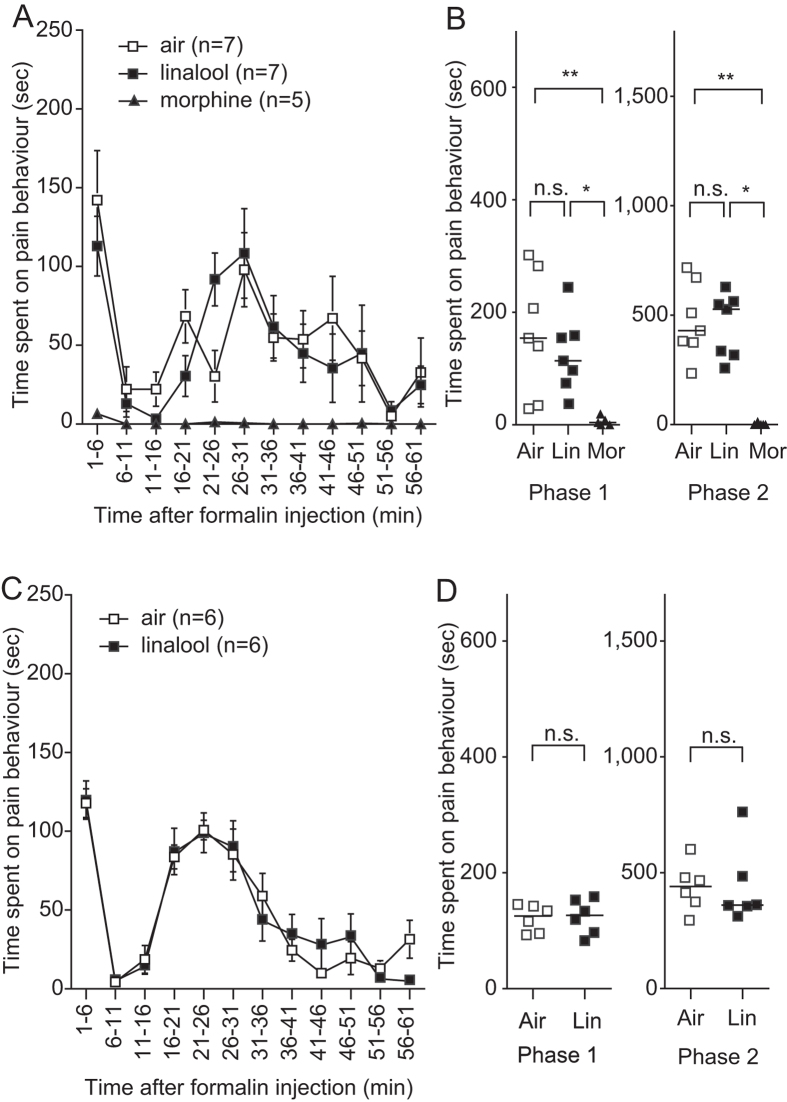
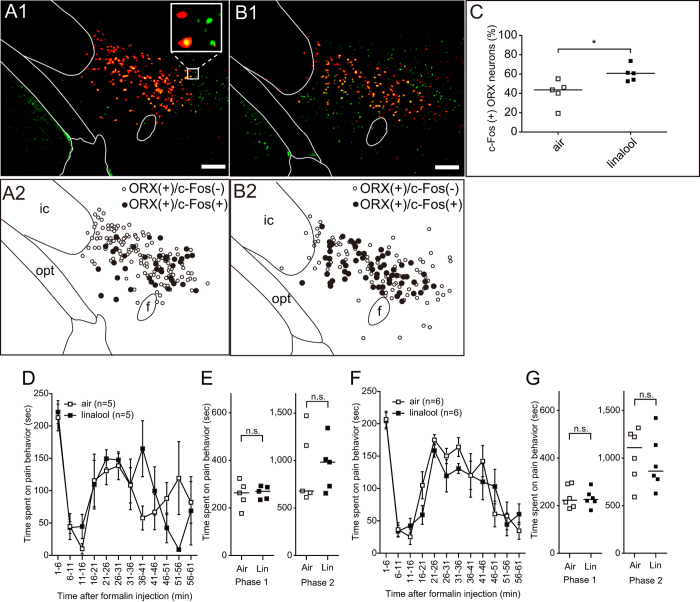
Various folk remedies employ certain odorous compounds with analgesic effects. In fact, linalool, a monoterpene alcohol found in lavender extracts, has been found to attenuate pain responses via subcutaneous, intraperitoneal, intrathecal, and oral administration. However, the analgesic effects of odorous compounds mediated by olfaction have not been thoroughly examined. We performed behavioural pain tests under odourant vapour exposure in mice. Among six odourant molecules examined, linalool significantly increased the pain threshold and attenuated pain behaviours. Olfactory bulb or epithelium lesion removed these effects, indicating that olfactory sensory input triggered the effects. Furthermore, immunohistochemical analysis revealed that linalool activated hypothalamic orexin neurons, one of the key mediators for pain processing. Formalin tests in orexin neuron-ablated and orexin peptide-deficient mice showed orexinergic transmission was essential for linalool odour-induced analgesia. Together, these findings reveal central analgesic circuits triggered by olfactory input in the mammalian brain and support a potential therapeutic approach for treating pain with linalool odour stimulation.
Figures
Similar articles
-
An anatomically distinct subpopulation of orexin neurons project from the lateral hypothalamus to the olfactory bulb.J Comp Neurol. 2023 Oct;531(15):1510-1524. doi: 10.1002/cne.25518. Epub 2023 Jul 11. J Comp Neurol. 2023. PMID: 37434469 Free PMC article.
-
Orexinergic descending inhibitory pathway mediates linalool odor-induced analgesia in mice.Sci Rep. 2021 Apr 29;11(1):9224. doi: 10.1038/s41598-021-88359-5. Sci Rep. 2021. PMID: 33927235 Free PMC article.
-
Analgesic effect of linalool odor on oral ulcerative mucositis-induced pain in rats.Brain Res Bull. 2024 Jan;206:110844. doi: 10.1016/j.brainresbull.2023.110844. Epub 2023 Dec 12. Brain Res Bull. 2024. PMID: 38096923
-
The circuits of the olfactory bulb. The exception as a rule.Anat Rec (Hoboken). 2013 Sep;296(9):1401-12. doi: 10.1002/ar.22732. Epub 2013 Jul 31. Anat Rec (Hoboken). 2013. PMID: 23907743 Review.
-
Hypothalamic modulation of breathing.Adv Exp Med Biol. 2010;669:243-7. doi: 10.1007/978-1-4419-5692-7_49. Adv Exp Med Biol. 2010. PMID: 20217358 Review.
Cited by
-
Possible Combinatorial Utilization of Phytochemicals and Extracellular Vesicles for Wound Healing and Regeneration.Int J Mol Sci. 2024 Sep 26;25(19):10353. doi: 10.3390/ijms251910353. Int J Mol Sci. 2024. PMID: 39408681 Free PMC article. Review.
-
Orexin Receptor Blockade-Induced Sleep Preserves the Ability to Wake in the Presence of Threat in Mice.Front Behav Neurosci. 2019 Jan 8;12:327. doi: 10.3389/fnbeh.2018.00327. eCollection 2018. Front Behav Neurosci. 2019. PMID: 30687033 Free PMC article.
-
Chemical Composition, Analgesic and Anti-Inflammatory Activity of Pelargonium peltatum Essential Oils from Eastern Cape, South Africa.Molecules. 2023 Jul 8;28(14):5294. doi: 10.3390/molecules28145294. Molecules. 2023. PMID: 37513168 Free PMC article.
-
The Mechanism of Compound Anshen Essential Oil in the Treatment of Insomnia Was Examined by Network Pharmacology.Evid Based Complement Alternat Med. 2019 Jun 4;2019:9241403. doi: 10.1155/2019/9241403. eCollection 2019. Evid Based Complement Alternat Med. 2019. PMID: 31275424 Free PMC article.
-
Analgesic Potential of Terpenes Derived from Cannabis sativa.Pharmacol Rev. 2021 Oct;73(4):98-126. doi: 10.1124/pharmrev.120.000046. Pharmacol Rev. 2021. PMID: 34663685 Free PMC article. Review.
References
-
- Peana A. T. et al.. (−)-Linalool produces antinociception in two experimental models of pain. Eur. J. Pharmacol. 460, 37–41 (2003). - PubMed
-
- Peana A. T. et al.. Profile of spinal and supra-spinal antinociception of (−)-linalool. Eur. J. Pharmacol. 485, 165–174 (2004). - PubMed
-
- Peana A. T. et al.. Involvement of adenosine A1 and A2A receptors in (−)-linalool-induced antinociception. Life Sci. 78, 2471–2474 (2006). - PubMed
-
- Batista P. A. et al.. The antinociceptive effect of (−)-linalool in models of chronic inflammatory and neuropathic hypersensitivity in mice. J. Pain 11, 1222–1229 (2010). - PubMed
-
- Batista P. A. et al.. Evidence for the involvement of ionotropic glutamatergic receptors on the antinociceptive effect of (−)-linalool in mice. Neurosci. Lett. 440, 299–303 (2008). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
