

TRIM28 multi-domain protein regulates cancer stem cell population in breast tumor development
- PMID: 27845900
- PMCID: PMC5352203
- DOI: 10.18632/oncotarget.13273
TRIM28 multi-domain protein regulates cancer stem cell population in breast tumor development
Abstract
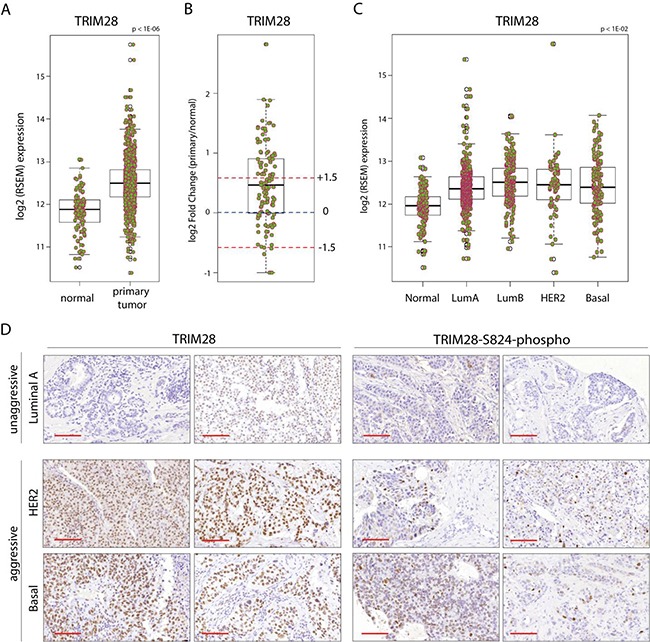
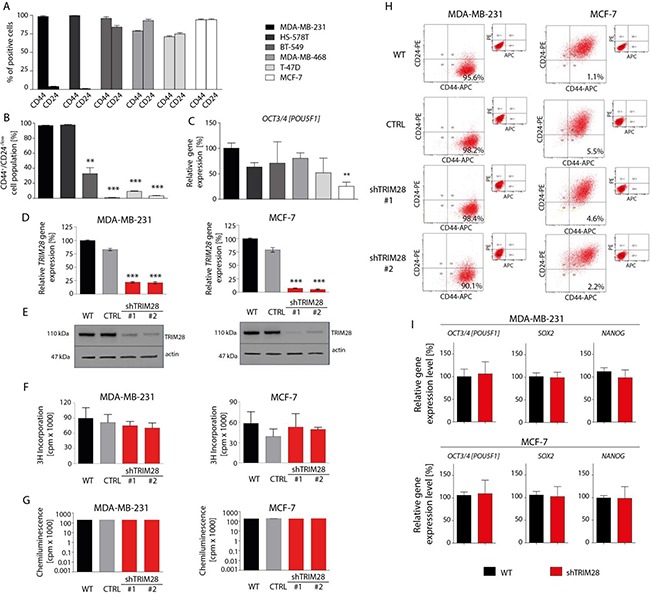
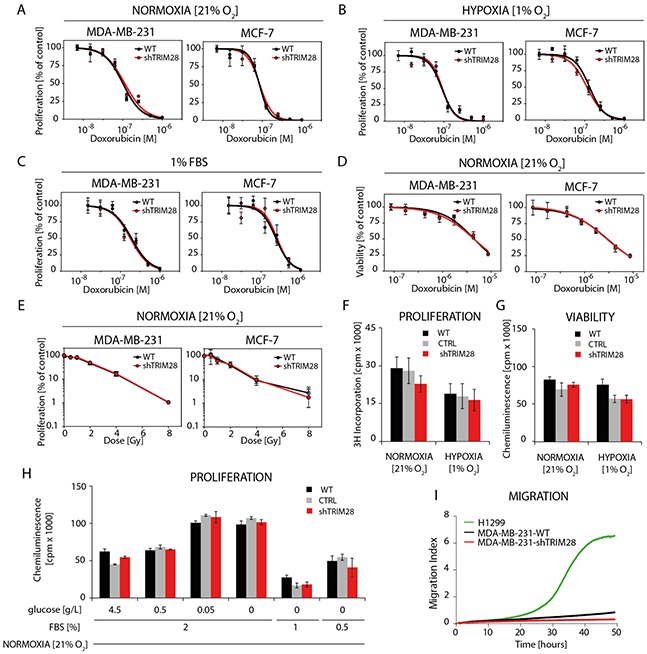
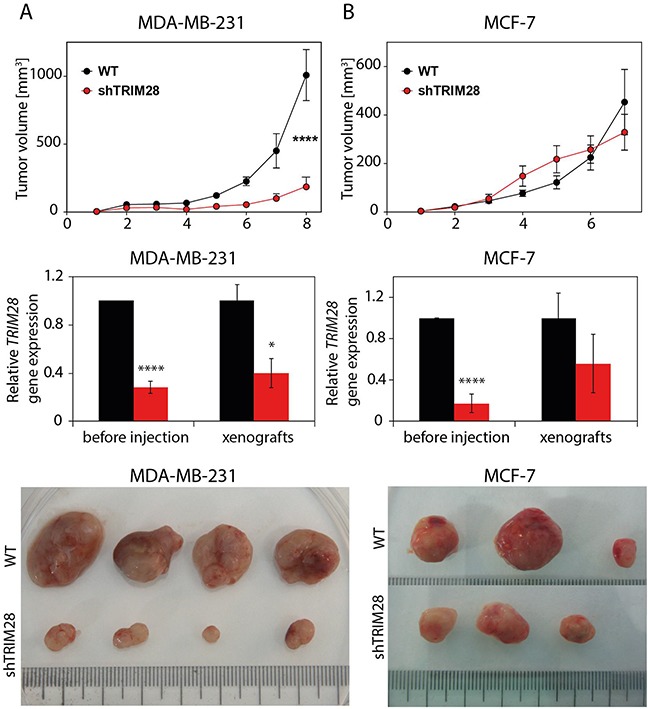
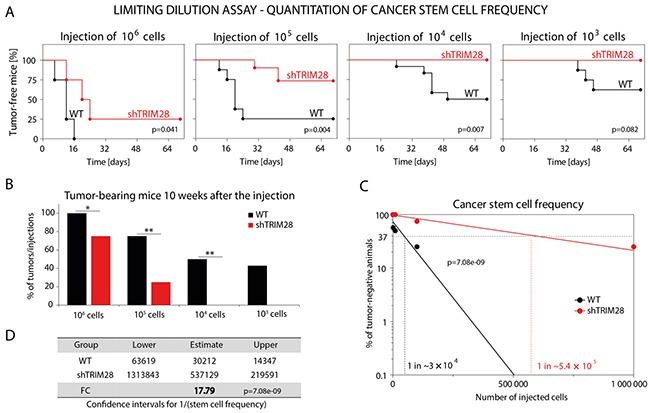
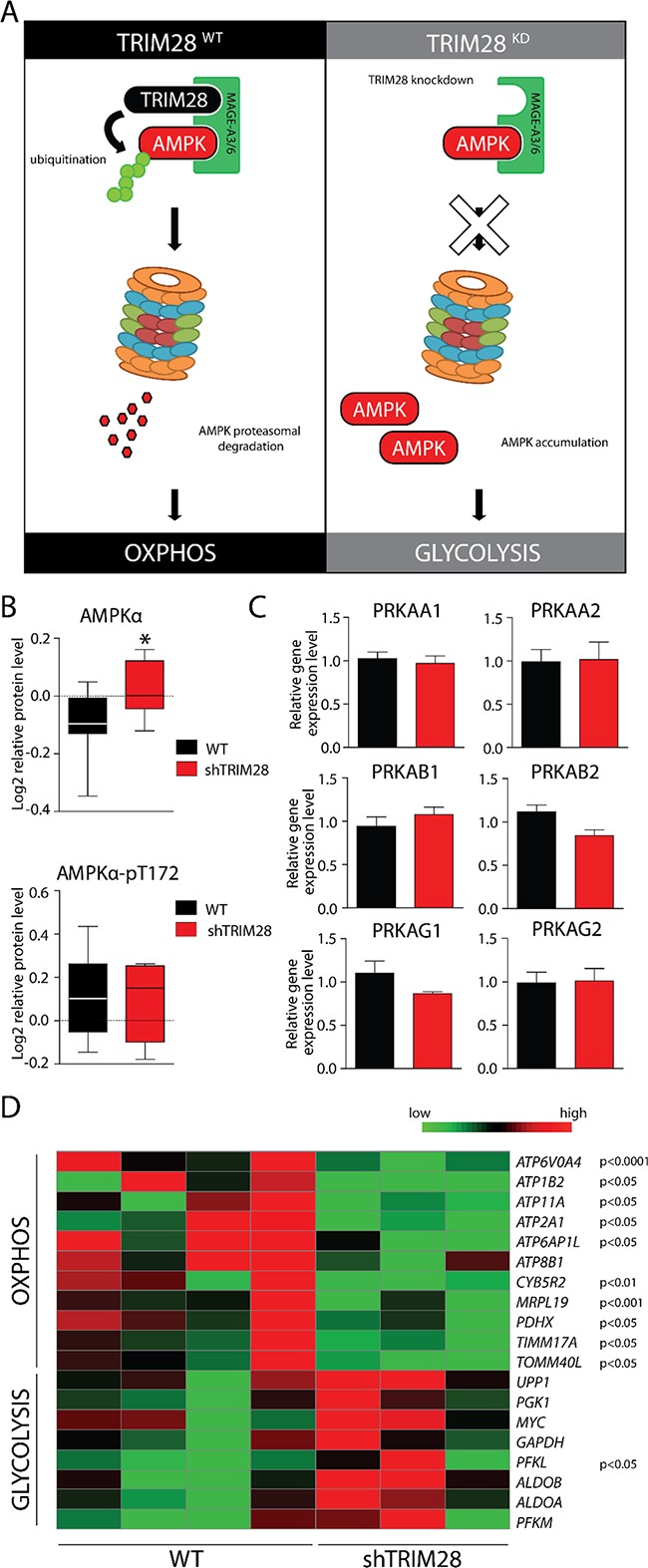
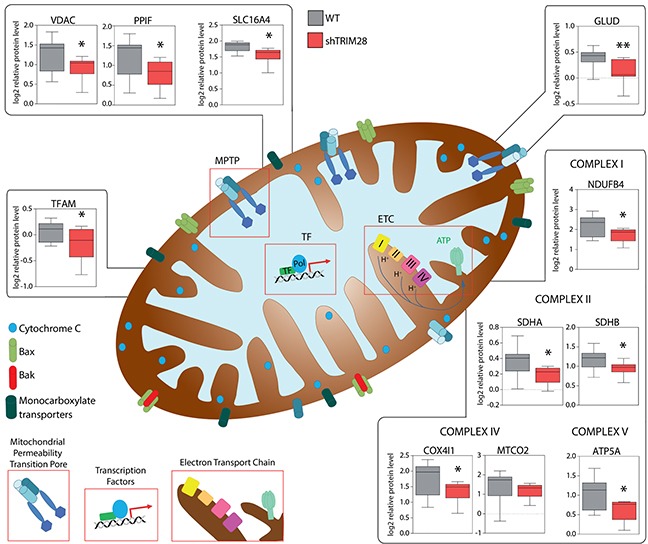
The expression of Tripartite motif-containing protein 28 (TRIM28)/Krüppel-associated box (KRAB)-associated protein 1 (KAP1), is elevated in at least 14 tumor types, including solid and hematopoietic tumors. High level of TRIM28 is associated with triple-negative subtype of breast cancer (TNBC), which shows higher aggressiveness and lower survival rates. Interestingly, TRIM28 is essential for maintaining the pluripotent phenotype in embryonic stem cells. Following on that finding, we evaluated the role of TRIM28 protein in the regulation of breast cancer stem cells (CSC) populations and tumorigenesis in vitro and in vivo. Downregulation of TRIM28 expression in xenografts led to deceased expression of pluripotency and mesenchymal markers, as well as inhibition of signaling pathways involved in the complex mechanism of CSC maintenance. Moreover, TRIM28 depletion reduced the ability of cancer cells to induce tumor growth when subcutaneously injected in limiting dilutions. Our data demonstrate that the downregulation of TRIM28 gene expression reduced the ability of CSCs to self-renew that resulted in significant reduction of tumor growth. Loss of function of TRIM28 leads to dysregulation of cell cycle, cellular response to stress, cancer cell metabolism, and inhibition of oxidative phosphorylation. All these mechanisms directly regulate maintenance of CSC population. Our original results revealed the role of the TRIM28 in regulating the CSC population in breast cancer. These findings may pave the way to novel and more effective therapies targeting cancer stem cells in breast tumors.
Keywords: KAP1; TRIM28; breast cancer stem cells; epigenetics; pluripotency.
Conflict of interest statement
The authors disclose no potential conflicts of interest.
Figures

Similar articles
-
Tripartite motif containing 28 (TRIM28) promotes breast cancer metastasis by stabilizing TWIST1 protein.Sci Rep. 2016 Jul 14;6:29822. doi: 10.1038/srep29822. Sci Rep. 2016. PMID: 27412325 Free PMC article.
-
Nestin positively regulates the Wnt/β-catenin pathway and the proliferation, survival and invasiveness of breast cancer stem cells.Breast Cancer Res. 2014 Jul 24;16(4):408. doi: 10.1186/s13058-014-0408-8. Breast Cancer Res. 2014. PMID: 25056574 Free PMC article.
-
Metalloprotease-disintegrin ADAM12 actively promotes the stem cell-like phenotype in claudin-low breast cancer.Mol Cancer. 2017 Feb 1;16(1):32. doi: 10.1186/s12943-017-0599-6. Mol Cancer. 2017. PMID: 28148288 Free PMC article.
-
The complexity of TRIM28 contribution to cancer.J Biomed Sci. 2017 Aug 29;24(1):63. doi: 10.1186/s12929-017-0374-4. J Biomed Sci. 2017. PMID: 28851455 Free PMC article. Review.
-
Developmental signaling pathways regulating mammary stem cells and contributing to the etiology of triple-negative breast cancer.Breast Cancer Res Treat. 2016 Apr;156(2):211-26. doi: 10.1007/s10549-016-3746-7. Epub 2016 Mar 11. Breast Cancer Res Treat. 2016. PMID: 26968398 Free PMC article. Review.
Cited by
-
The Association between TIF1 Family Members and Cancer Stemness in Solid Tumors.Cancers (Basel). 2021 Mar 26;13(7):1528. doi: 10.3390/cancers13071528. Cancers (Basel). 2021. PMID: 33810347 Free PMC article.
-
Disruption of RING and PHD Domains of TRIM28 Evokes Differentiation in Human iPSCs.Cells. 2021 Jul 29;10(8):1933. doi: 10.3390/cells10081933. Cells. 2021. PMID: 34440702 Free PMC article.
-
KAP1 promotes gastric adenocarcinoma progression by activating Hippo/YAP1 signaling via binding to HNRNPAB.Cancer Lett. 2025 Jul 1;621:217695. doi: 10.1016/j.canlet.2025.217695. Epub 2025 Apr 4. Cancer Lett. 2025. PMID: 40189014 Free PMC article.
-
Mapping of the contraction-induced phosphoproteome identifies TRIM28 as a significant regulator of skeletal muscle size and function.Cell Rep. 2021 Mar 2;34(9):108796. doi: 10.1016/j.celrep.2021.108796. Cell Rep. 2021. PMID: 33657380 Free PMC article.
-
The role of TRIM family proteins in the regulation of cancer stem cell self-renewal.Stem Cells. 2020 Feb;38(2):165-173. doi: 10.1002/stem.3109. Epub 2019 Nov 9. Stem Cells. 2020. PMID: 31664748 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
