

Ubiquitination of ERMES components by the E3 ligase Rsp5 is involved in mitophagy
- PMID: 27846375
- PMCID: PMC5240830
- DOI: 10.1080/15548627.2016.1252889
Ubiquitination of ERMES components by the E3 ligase Rsp5 is involved in mitophagy
Abstract
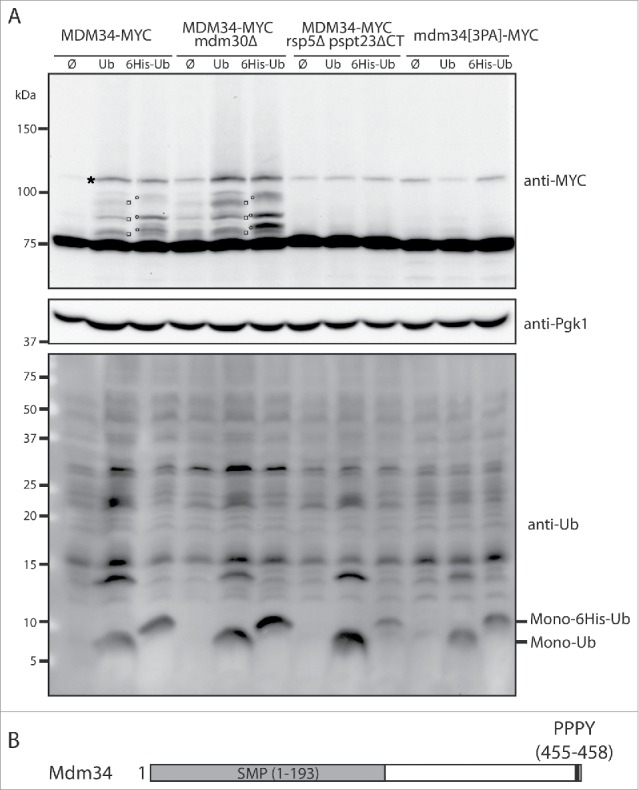
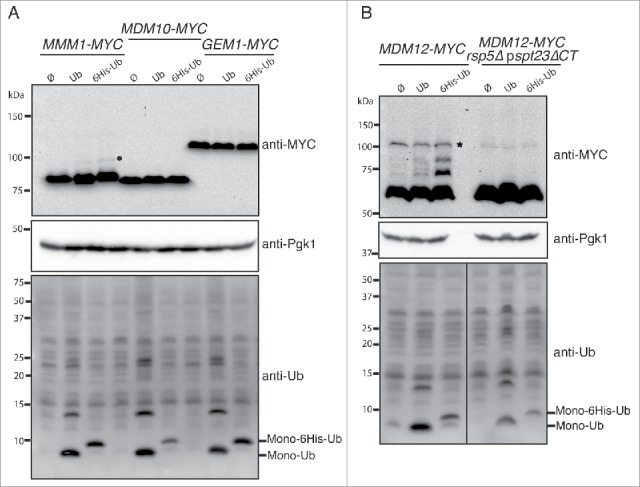
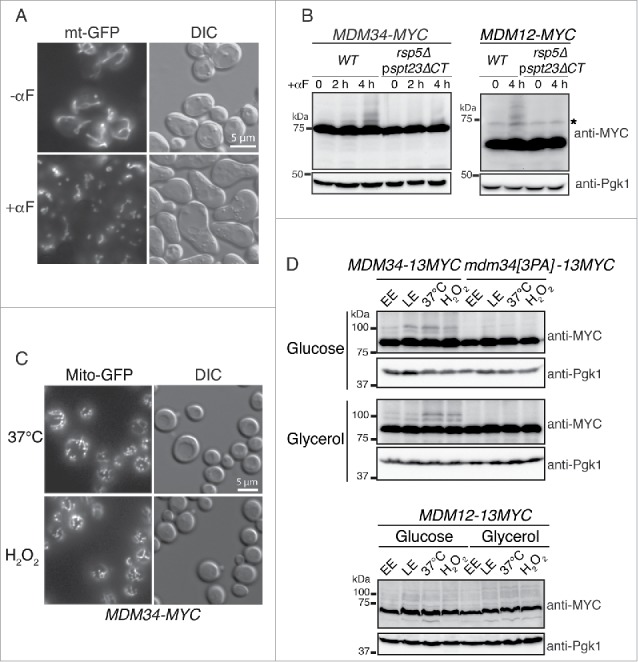
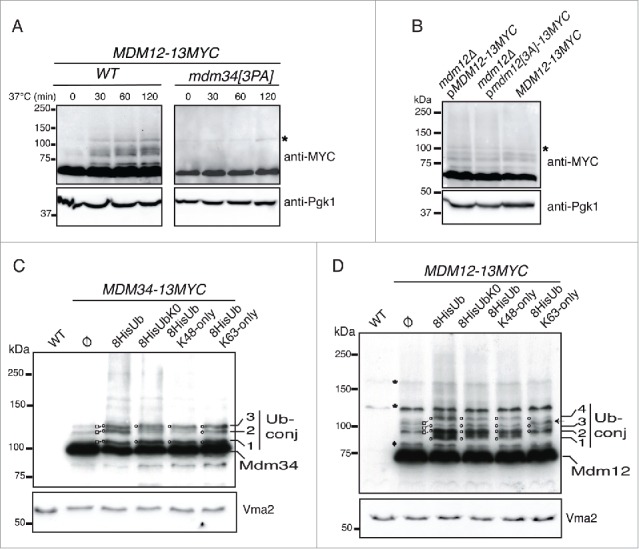
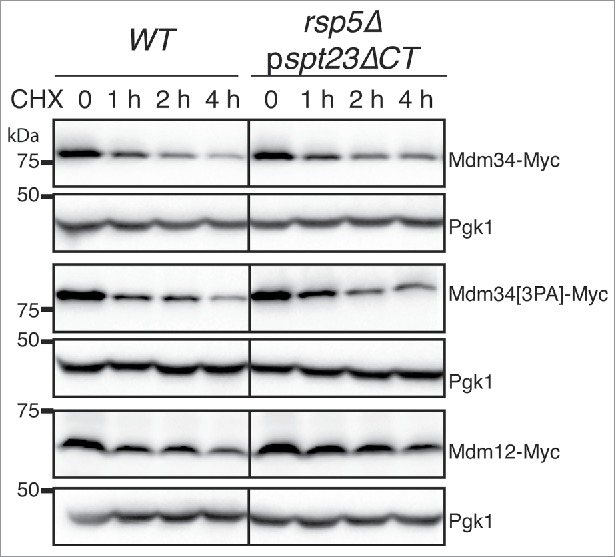
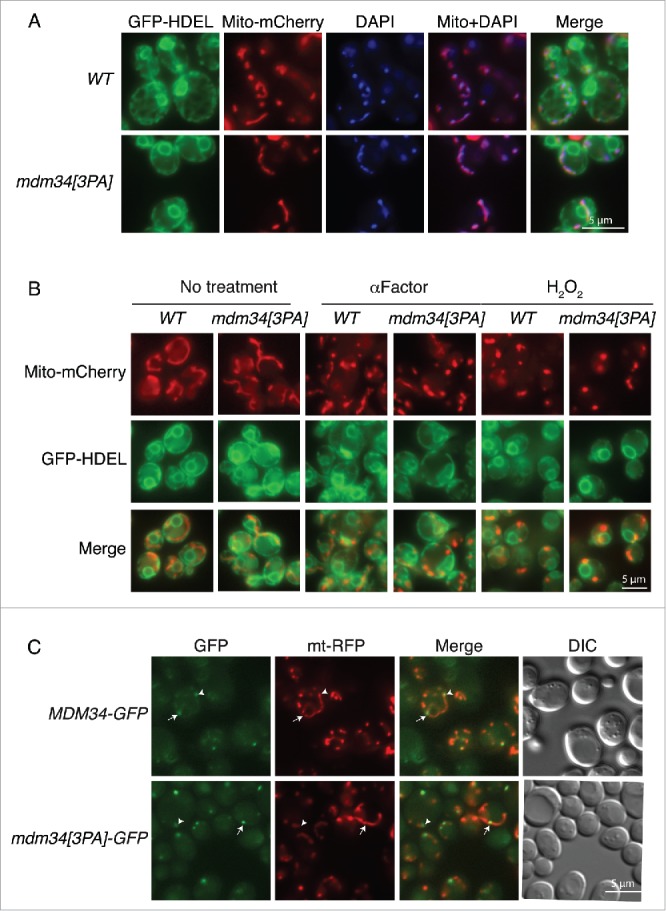
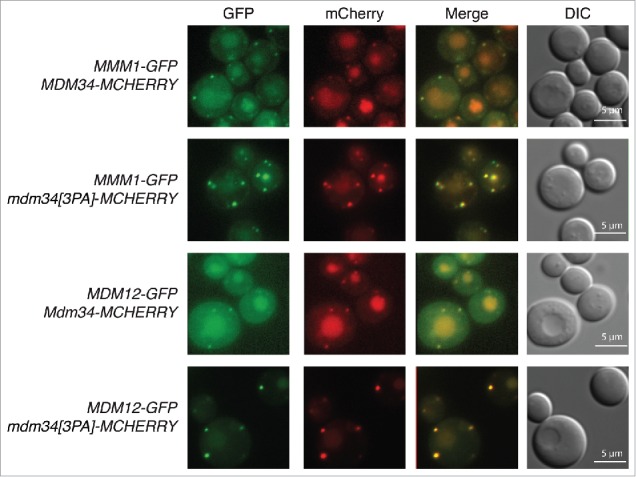
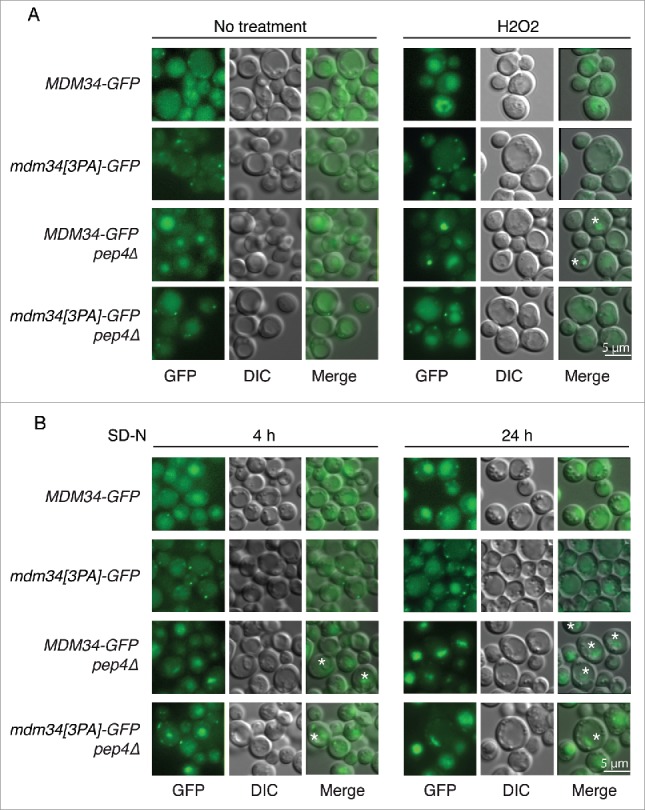
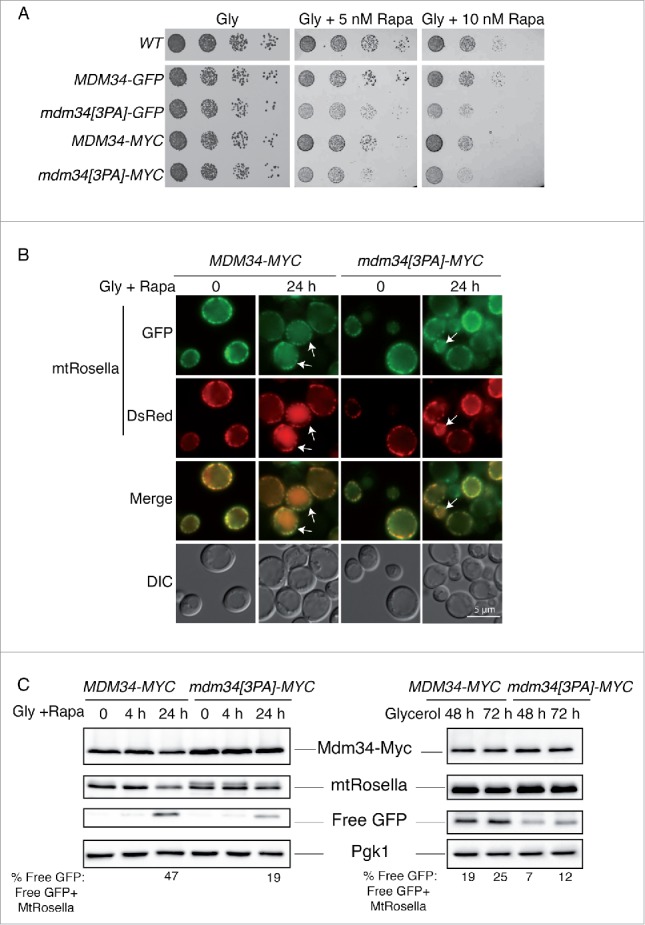
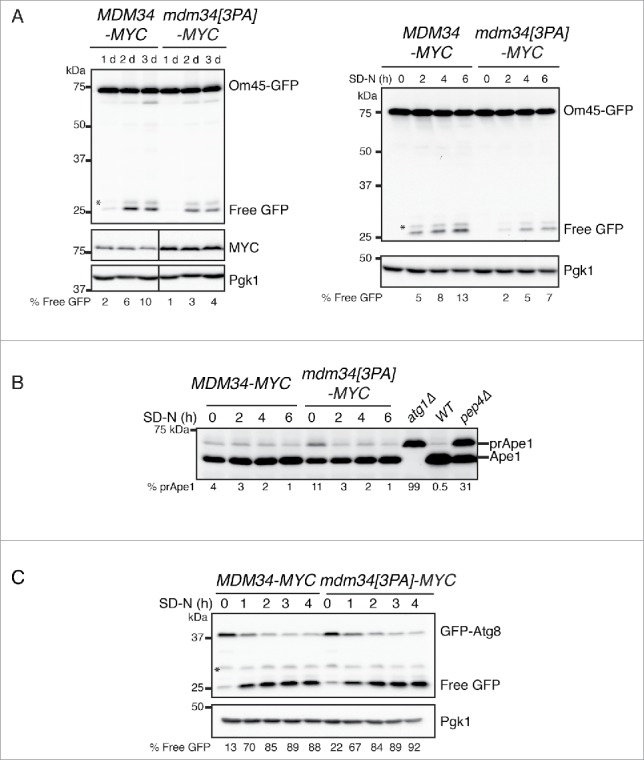
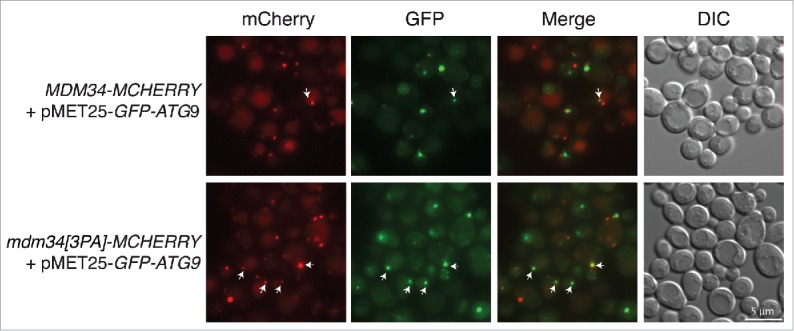
Mitochondria are dynamic organelles that undergo permanent fission and fusion events. These processes play an essential role in maintaining normal cellular function. In the yeast Saccharomyces cerevisiae, the endoplasmic reticulum-mitochondrial encounter structure (ERMES) is a marker of sites of mitochondrial division, but it is also involved in a plethora of other mitochondrial functions. However, it remains unclear how these different functions are regulated. We show here that Mdm34 and Mdm12, 2 components of ERMES, are ubiquitinated by the E3 ligase Rsp5. This ubiquitination is not involved in mitochondrial dynamics or in the distribution and turnover of ERMES. Nevertheless, the ubiquitination of Mdm34 and Mdm12 was required for efficient mitophagy. We thus report here the first identification of ubiquitinated substrates participating in yeast mitophagy.
Keywords: ER; ERMES; Mdm12; Mdm34; Rsp5; S. cerevisiae; mitochondria; mitophagy; ubiquitin.
Figures
References
-
- Westermann B. Mitochondrial fusion and fission in cell life and death. Nat Rev Mol Cell Biol 2010; 11:872-84; PMID:21102612; http://dx.doi.org/ 10.1038/nrm3013 - DOI - PubMed
-
- Mishra P, Chan DC. Mitochondrial dynamics and inheritance during cell division, development and disease. Nat Rev Mol Cell Biol 2014; 15:634-46; PMID:25237825; http://dx.doi.org/ 10.1038/nrm3877 - DOI - PMC - PubMed
-
- Friedman JR, Nunnari J. Mitochondrial form and function. Nature 2014; 505:335-43; PMID:24429632; http://dx.doi.org/ 10.1038/nature12985 - DOI - PMC - PubMed
-
- Friedman JR, Lackner LL, West M, Dibenedetto JR, Nunnari J, Voeltz GK. ER Tubules mark sites of mitochondrial division. Science 2011; 334:358-62; PMID:21885730; http://dx.doi.org/ 10.1126/science.1207385 - DOI - PMC - PubMed
-
- Klecker T, Böckler S, Westermann B. Making connections: interorganelle contacts orchestrate mitochondrial behavior. Trends Cell Biol 2014:1-9 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases