

Extensive migration of young neurons into the infant human frontal lobe
- PMID: 27846470
- PMCID: PMC5436574
- DOI: 10.1126/science.aaf7073
Extensive migration of young neurons into the infant human frontal lobe
Abstract
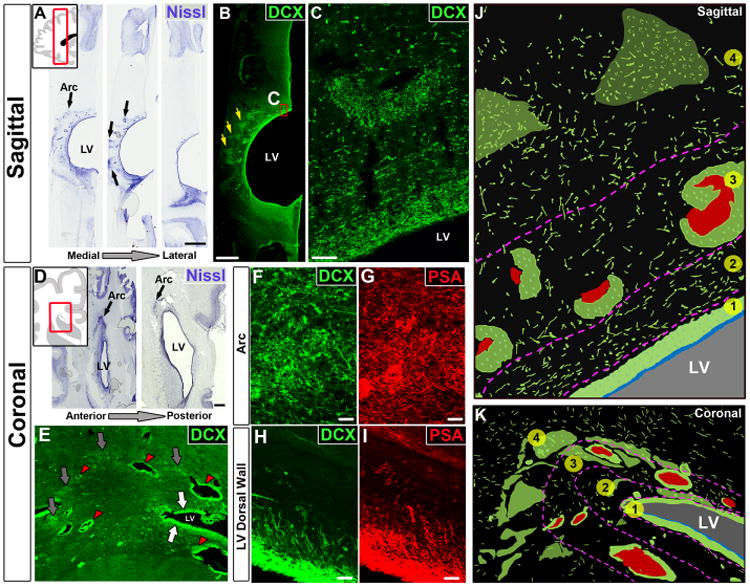
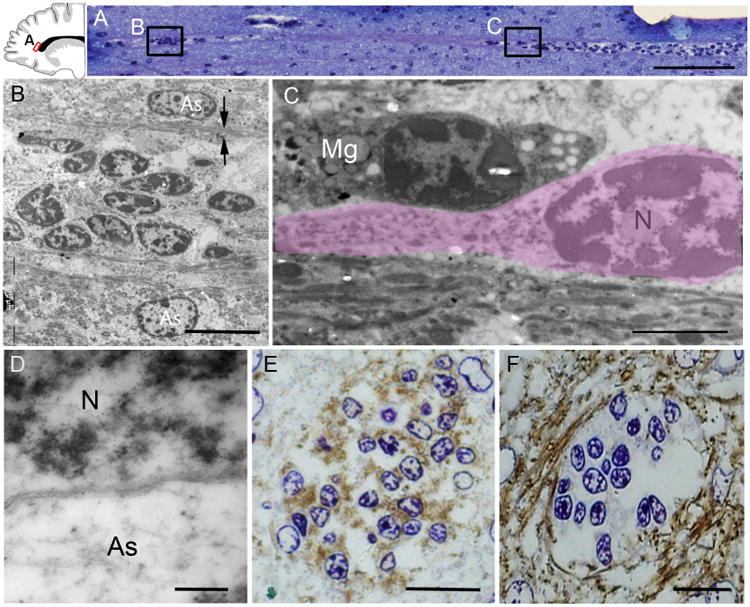
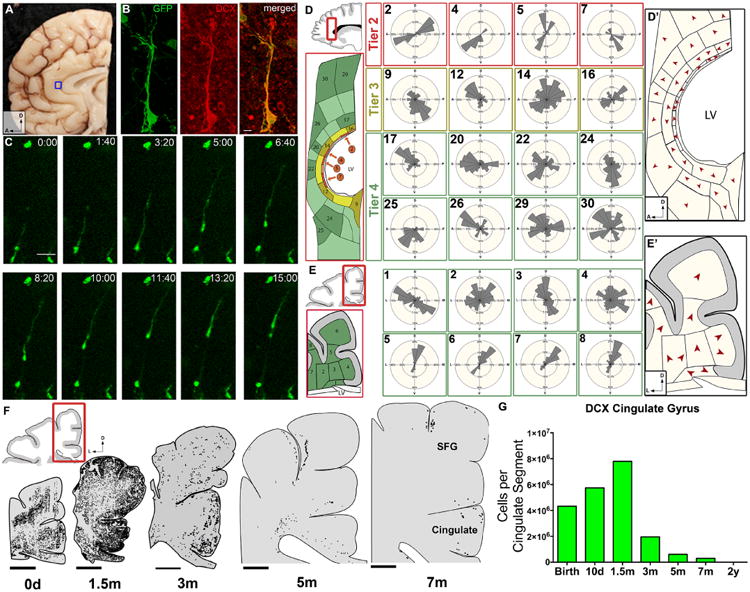
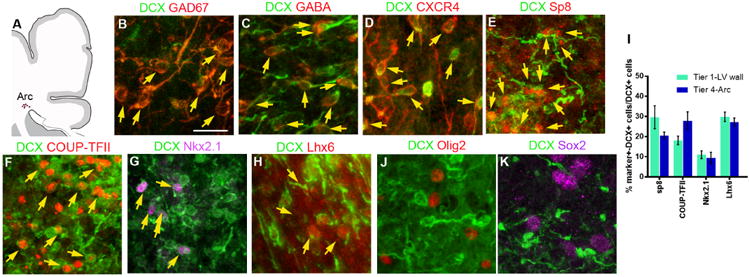
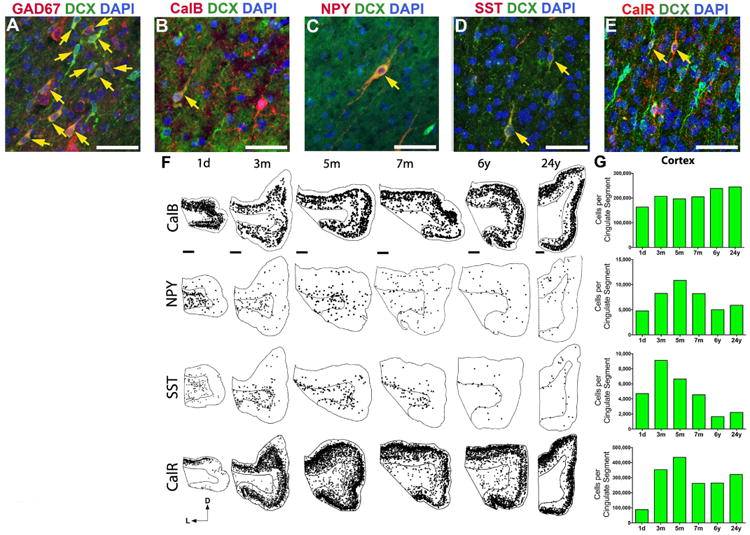
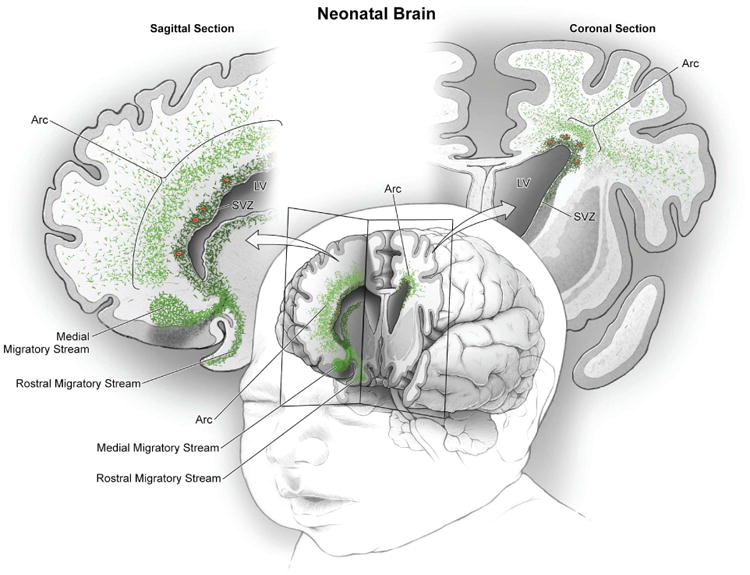
The first few months after birth, when a child begins to interact with the environment, are critical to human brain development. The human frontal lobe is important for social behavior and executive function; it has increased in size and complexity relative to other species, but the processes that have contributed to this expansion are unknown. Our studies of postmortem infant human brains revealed a collection of neurons that migrate and integrate widely into the frontal lobe during infancy. Chains of young neurons move tangentially close to the walls of the lateral ventricles and along blood vessels. These cells then individually disperse long distances to reach cortical tissue, where they differentiate and contribute to inhibitory circuits. Late-arriving interneurons could contribute to developmental plasticity, and the disruption of their postnatal migration or differentiation may underlie neurodevelopmental disorders.
Copyright © 2016, American Association for the Advancement of Science.
Figures
Comment in
-
Human brains teach us a surprising lesson.Science. 2016 Oct 7;354(6308):38-39. doi: 10.1126/science.aai9379. Science. 2016. PMID: 27846485 No abstract available.
-
Letter: Extensive Migration of Young Neurons Into the Infant Human Frontal Lobe.Neurosurgery. 2017 Aug 1;81(2):E16-E18. doi: 10.1093/neuros/nyx202. Neurosurgery. 2017. PMID: 28431048 Free PMC article. No abstract available.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
