

Molecular force spectroscopy with a DNA origami-based nanoscopic force clamp
- PMID: 27846560
- PMCID: PMC6546592
- DOI: 10.1126/science.aah5974
Molecular force spectroscopy with a DNA origami-based nanoscopic force clamp
Abstract
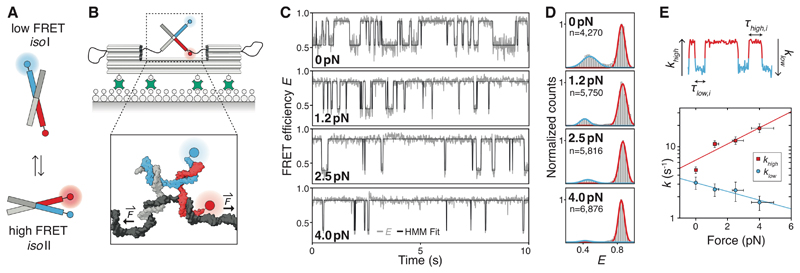
Forces in biological systems are typically investigated at the single-molecule level with atomic force microscopy or optical and magnetic tweezers, but these techniques suffer from limited data throughput and their requirement for a physical connection to the macroscopic world. We introduce a self-assembled nanoscopic force clamp built from DNA that operates autonomously and allows massive parallelization. Single-stranded DNA sections of an origami structure acted as entropic springs and exerted controlled tension in the low piconewton range on a molecular system, whose conformational transitions were monitored by single-molecule Förster resonance energy transfer. We used the conformer switching of a Holliday junction as a benchmark and studied the TATA-binding protein-induced bending of a DNA duplex under tension. The observed suppression of bending above 10 piconewtons provides further evidence of mechanosensitivity in gene regulation.
Copyright © 2016, American Association for the Advancement of Science.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
-
- Kellermayer MS, Smith SB, Granzier HL, Bustamante C. Folding-unfolding transitions in single titin molecules characterized with laser tweezers. Science. 1997;276:1112–1116. - PubMed
-
- Rief M, Gautel M, Oesterhelt F, Fernandez JM, Gaub HE. Reversible unfolding of individual titin immunoglobulin domains by AFM. Science. 1997;276:1109–1112. - PubMed
-
- Smith SB, Cui Y, Bustamante C. Overstretching B-DNA: the elastic response of individual double-stranded and single-stranded DNA molecules. Science. 1996;271:795–799. - PubMed
-
- Fernandez JM, Li H. Force-clamp spectroscopy monitors the folding trajectory of a single protein. Science. 2004;303:1674–1678. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
