

Modulation of NMDA channel gating by Ca2+ and Cd2+ binding to the external pore mouth
- PMID: 27848984
- PMCID: PMC5111045
- DOI: 10.1038/srep37029
Modulation of NMDA channel gating by Ca2+ and Cd2+ binding to the external pore mouth
Abstract
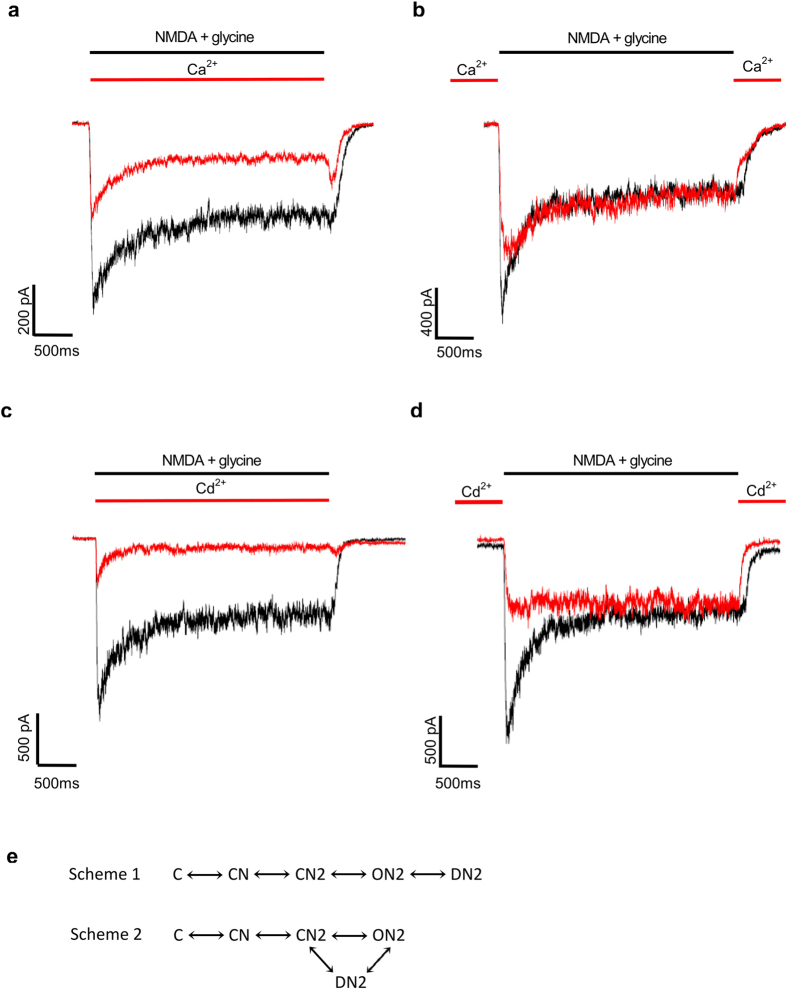
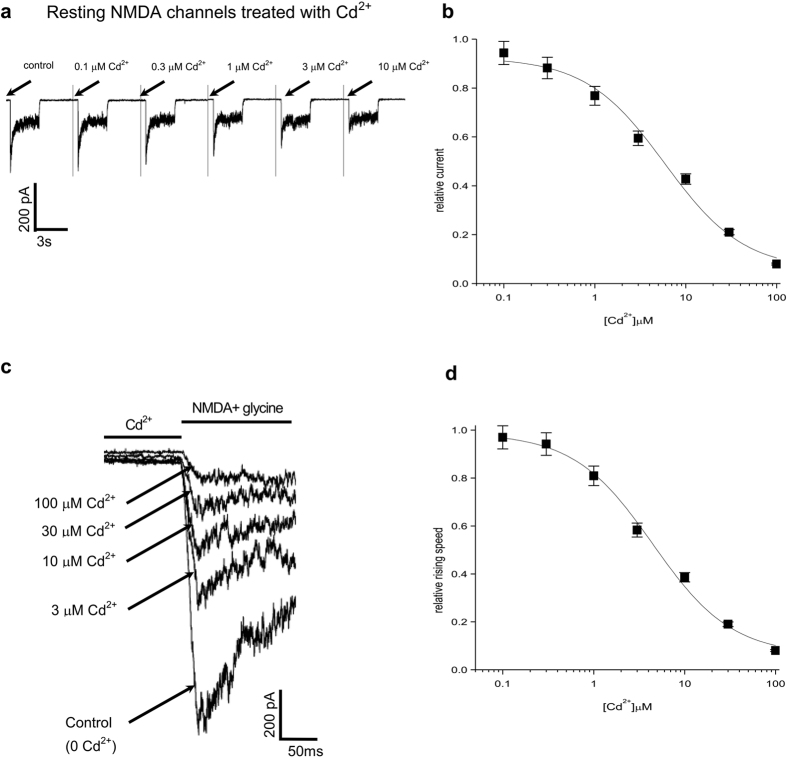
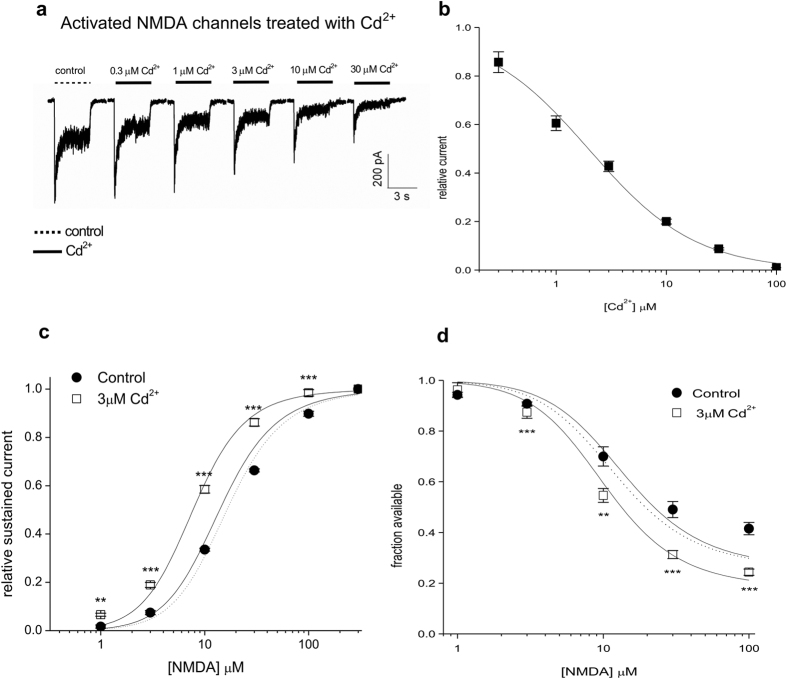
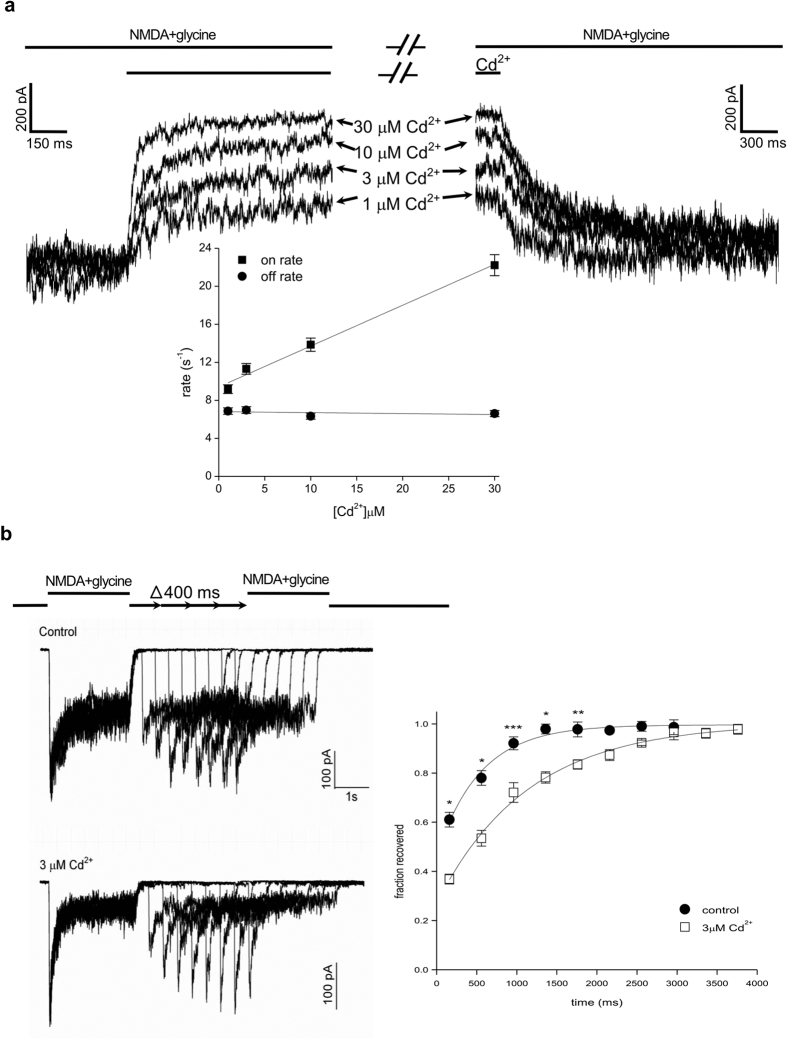
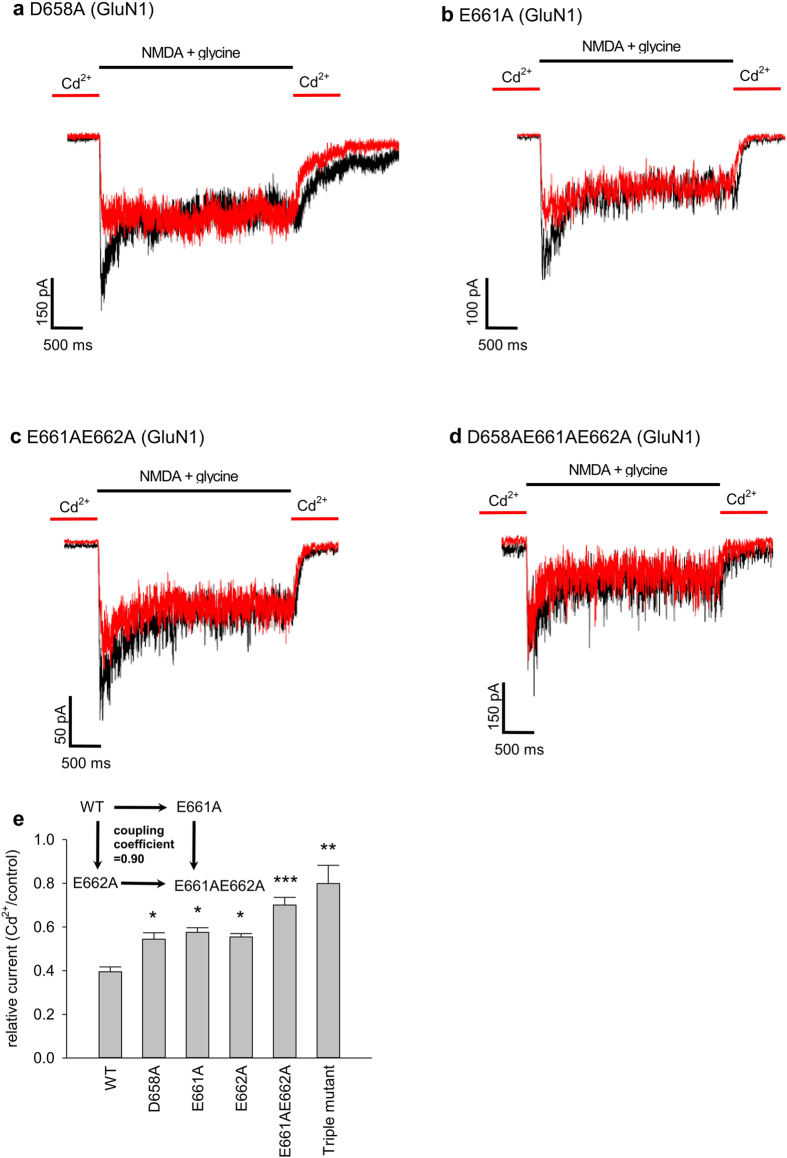
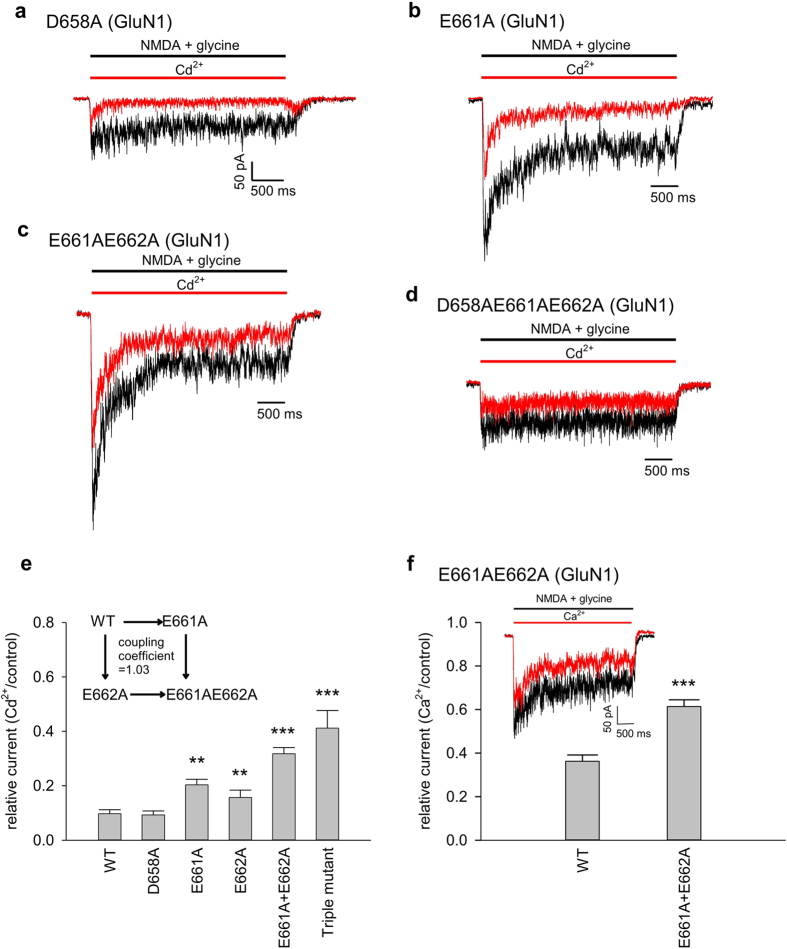
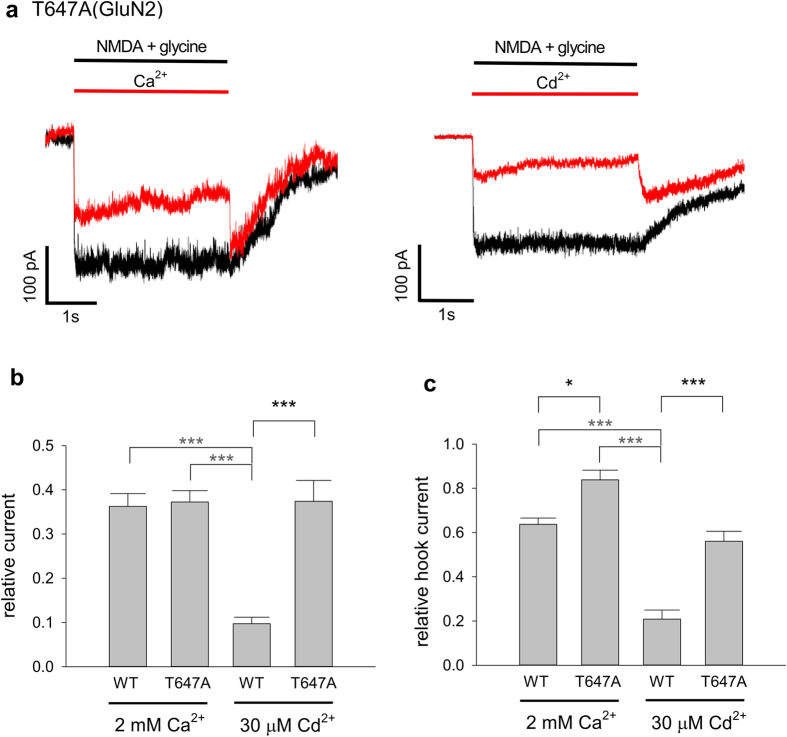
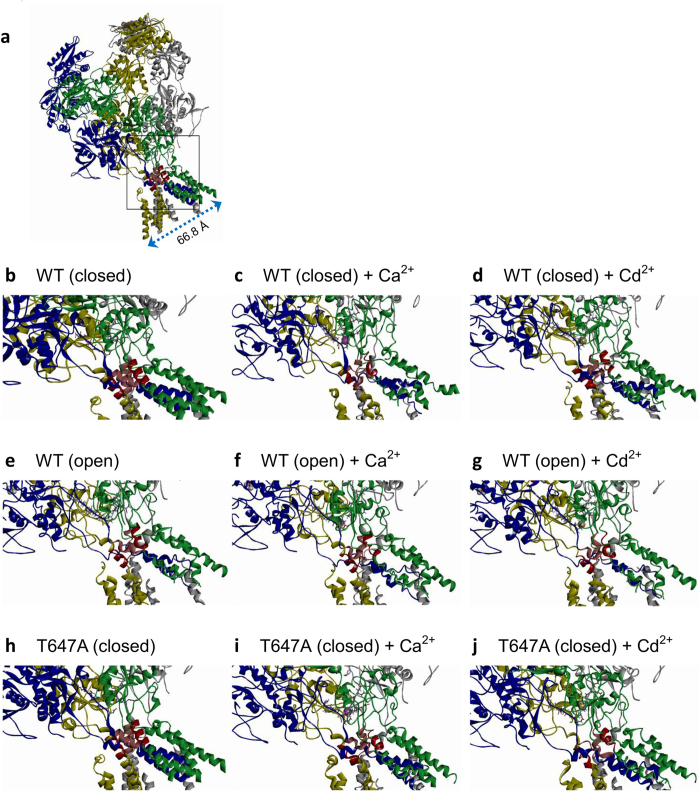
NMDA receptor channels are characterized by high Ca2+ permeability. It remains unclear whether extracellular Ca2+ could directly modulate channel gating and control Ca2+ influxes. We demonstrate a pore-blocking site external to the activation gate for extracellular Ca2+ and Cd2+, which has the same charge and radius as Ca2+ but is impermeable to the channel. The apparent affinity of Cd2+ or Ca2+ is higher toward the activated (a steady-state mixture of the open and desensitized, probably chiefly the latter) than the closed states. The blocking effect of Cd2+ is well correlated with the number of charges in the DRPEER motif at the external pore mouth, with coupling coefficients close to 1 in double mutant cycle analyses. The effect of Ca2+ and especially Cd2+ could be allosterically affected by T647A mutation located just inside the activation gate. A prominent "hook" also develops after wash-off of Cd2+ or Ca2+, suggesting faster unbinding rates of Cd2+ and Ca2+ with the mutation. We conclude that extracellular Ca2+ or Cd2+ directly binds to the DRPEER motif to modify NMDA channel activation (opening as well as desensitization), which seems to involve essential regional conformational changes centered at the bundle crossing point A652 (GluN1)/A651(GluN2).
Figures
References
-
- Wisden W. & Seeburg P. H. Mammalian ionotropic glutamate receptors. Curr Opin Neurobiol 3(3), 291–298 (1993). - PubMed
-
- Schoepfer. R. et al. Molecular biology of glutamate receptors. Prog Neurobiol 42(2), 353–357 (1994). - PubMed
-
- Gloor P. Epilepsy: relationships between electrophysiology and intracellular mechanisms involving second messengers and gene expression. Can J Neurol Sci. 16(1), 8–21 (1989). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
