

RNA modification enzyme TruB is a tRNA chaperone
- PMID: 27849601
- PMCID: PMC5167154
- DOI: 10.1073/pnas.1607512113
RNA modification enzyme TruB is a tRNA chaperone
Abstract
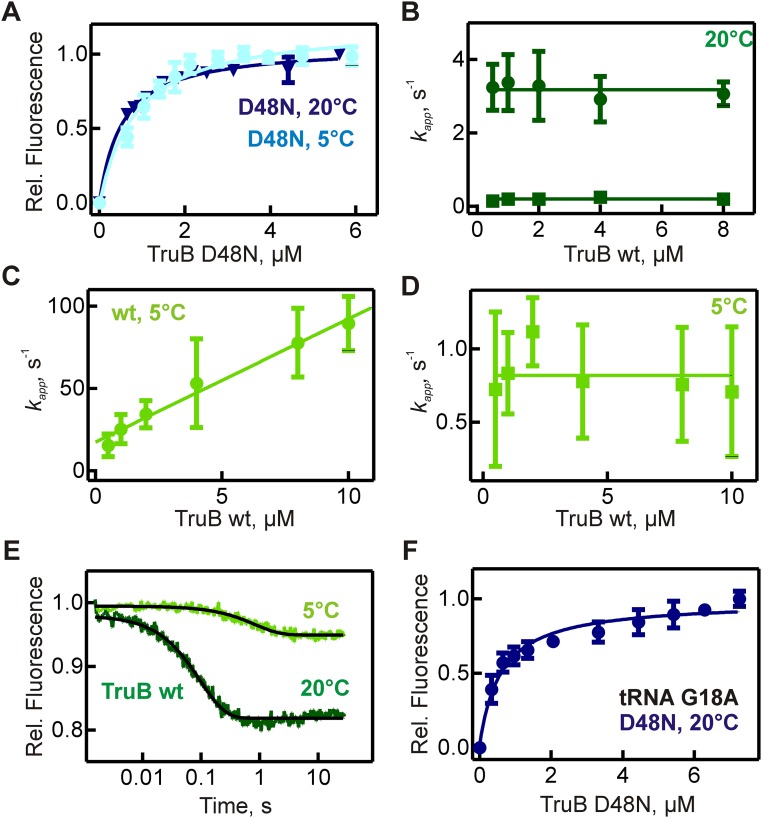
Cellular RNAs are chemically modified by many RNA modification enzymes; however, often the functions of modifications remain unclear, such as for pseudouridine formation in the tRNA TΨC arm by the bacterial tRNA pseudouridine synthase TruB. Here we test the hypothesis that RNA modification enzymes also act as RNA chaperones. Using TruB as a model, we demonstrate that TruB folds tRNA independent of its catalytic activity, thus increasing the fraction of tRNA that can be aminoacylated. By rapid kinetic stopped-flow analysis, we identified the molecular mechanism of TruB's RNA chaperone activity: TruB binds and unfolds both misfolded and folded tRNAs thereby providing misfolded tRNAs a second chance at folding. Previously, it has been shown that a catalytically inactive TruB variant has no phenotype when expressed in an Escherichia coli truB KO strain [Gutgsell N, et al. (2000) RNA 6(12):1870-1881]. However, here we uncover that E. coli strains expressing a TruB variant impaired in tRNA binding and in in vitro tRNA folding cannot compete with WT E. coli. Consequently, the tRNA chaperone activity of TruB is critical for bacterial fitness. In conclusion, we prove the tRNA chaperone activity of the pseudouridine synthase TruB, reveal its molecular mechanism, and demonstrate its importance for cellular fitness. We discuss the likelihood that other RNA modification enzymes are also RNA chaperones.
Keywords: RNA chaperone; RNA folding; RNA modification; pseudouridine; tRNA.
Conflict of interest statement
The authors declare no conflict of interest.
Figures












Comment in
-
Two for the price of one: RNA modification enzymes as chaperones.Proc Natl Acad Sci U S A. 2016 Dec 13;113(50):14176-14178. doi: 10.1073/pnas.1617402113. Epub 2016 Nov 30. Proc Natl Acad Sci U S A. 2016. PMID: 27911836 Free PMC article. No abstract available.
References
-
- Rajkowitsch L, et al. RNA chaperones, RNA annealers and RNA helicases. RNA Biol. 2007;4(3):118–130. - PubMed
-
- Ishitani R, Yokoyama S, Nureki O. Structure, dynamics, and function of RNA modification enzymes. Curr Opin Struct Biol. 2008;18(3):330–339. - PubMed
-
- Charette M, Gray MW. Pseudouridine in RNA: What, where, how, and why. IUBMB Life. 2000;49(5):341–351. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
