

Genetic and mechanistic diversity of piRNA 3'-end formation
- PMID: 27851737
- PMCID: PMC5164936
- DOI: 10.1038/nature20162
Genetic and mechanistic diversity of piRNA 3'-end formation
Abstract
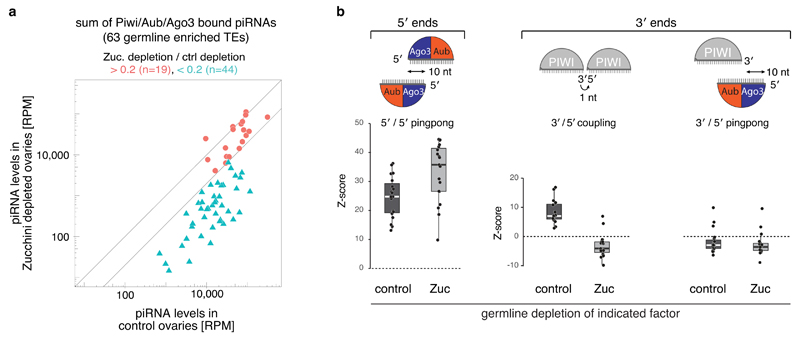
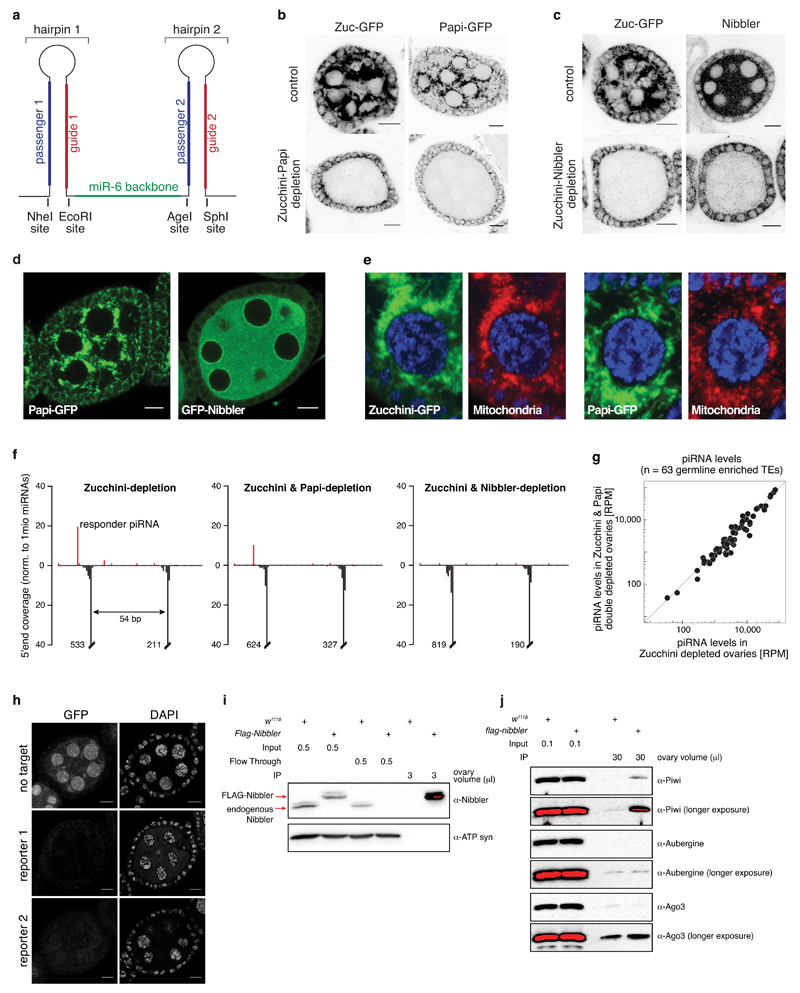
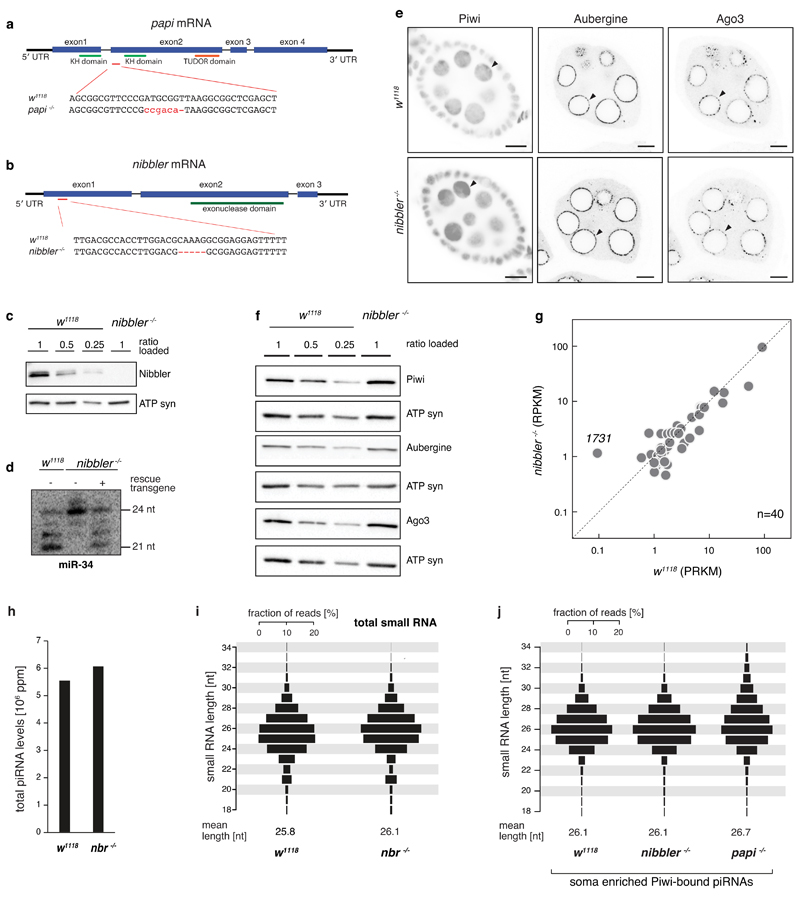
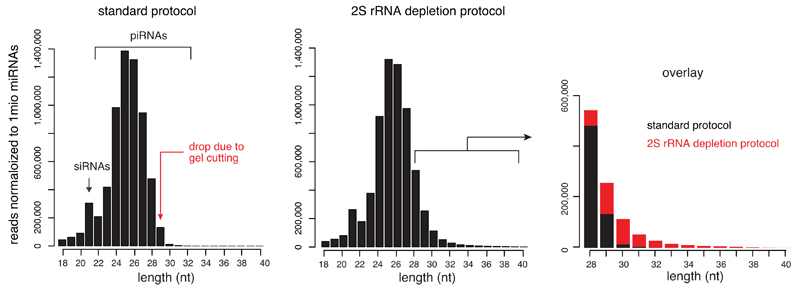
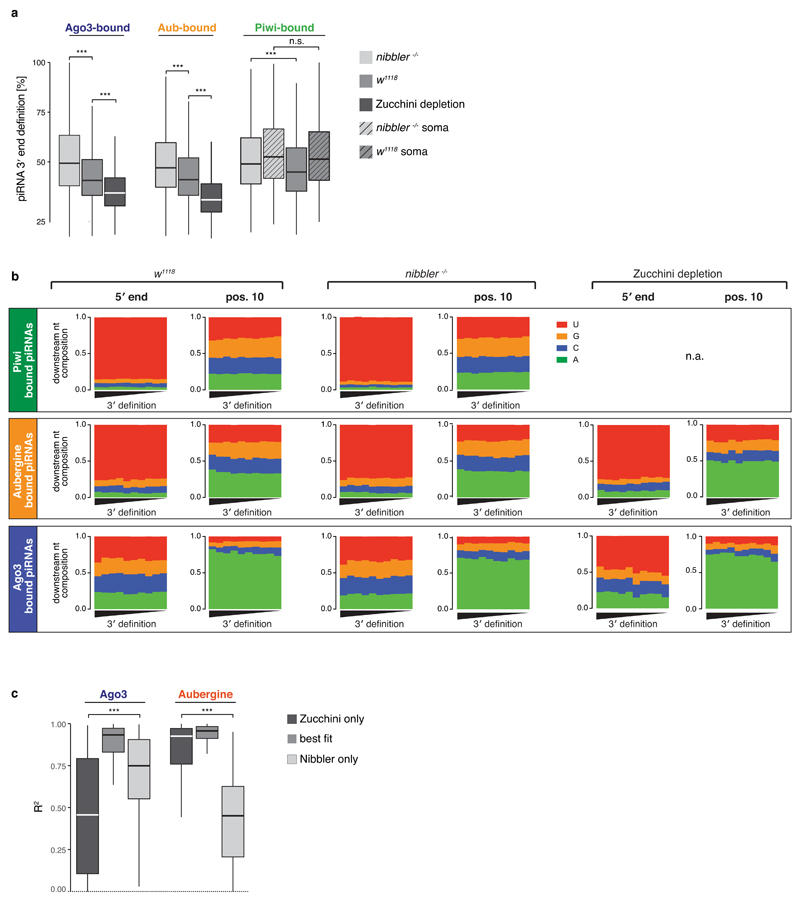
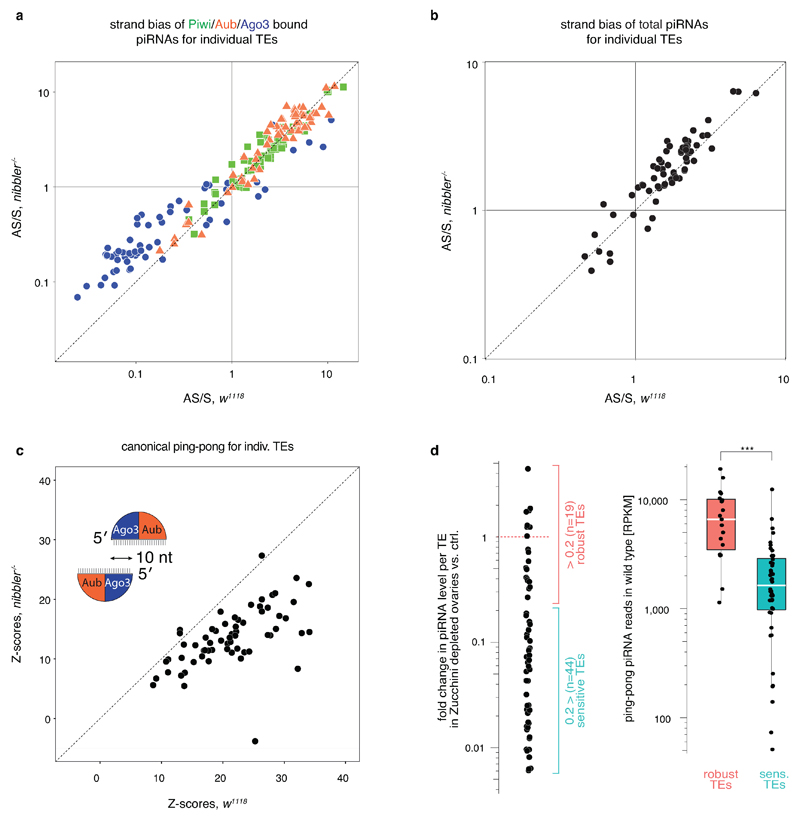
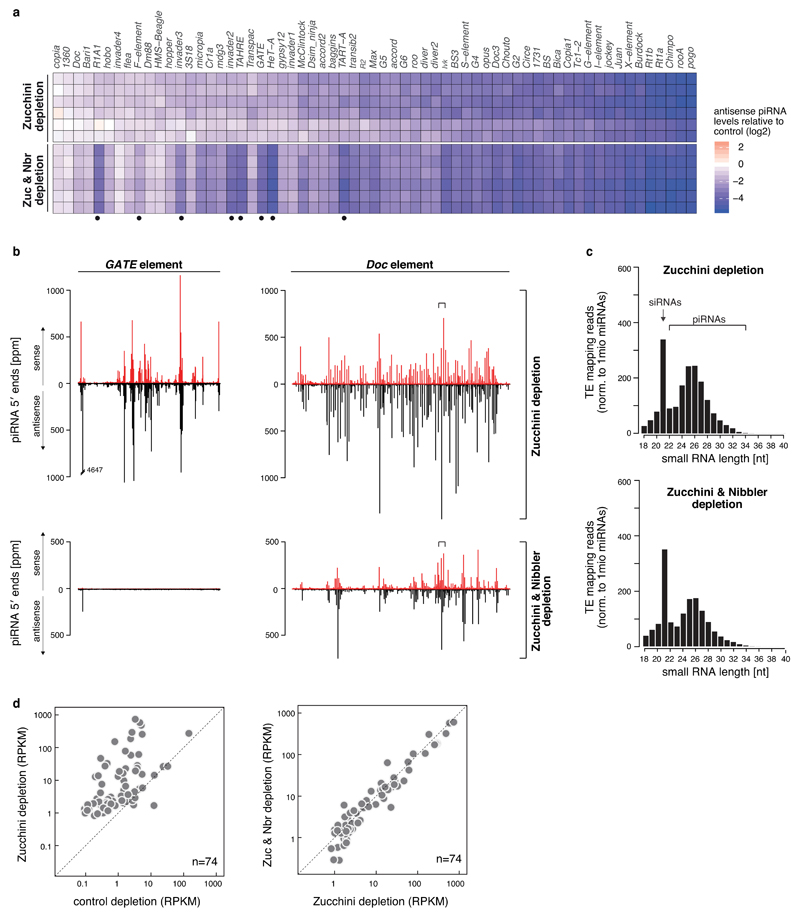
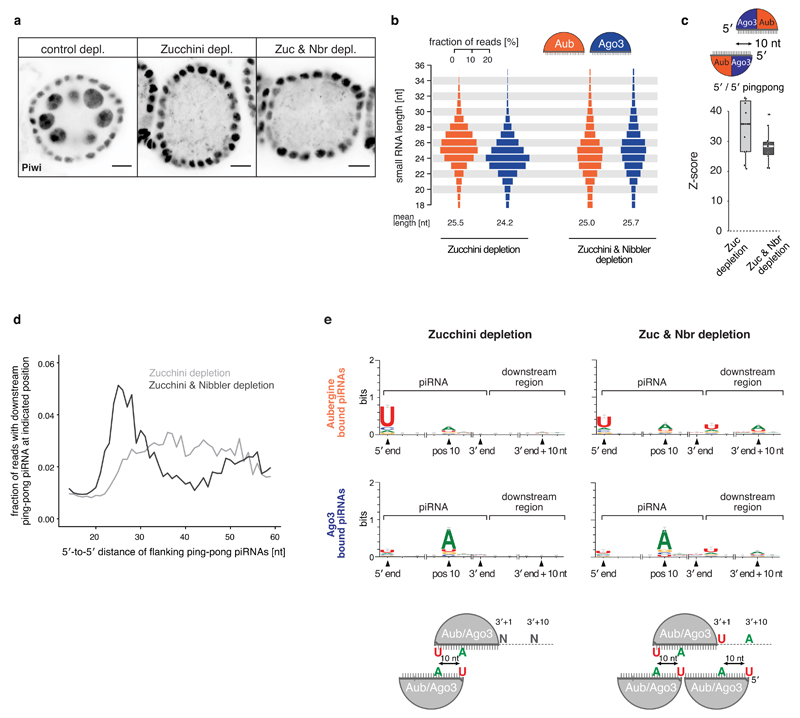
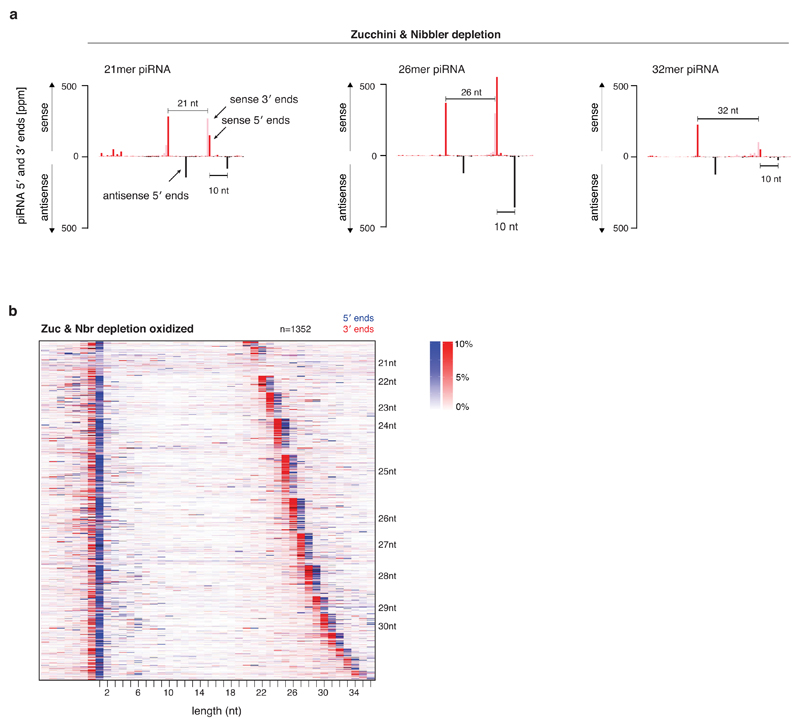
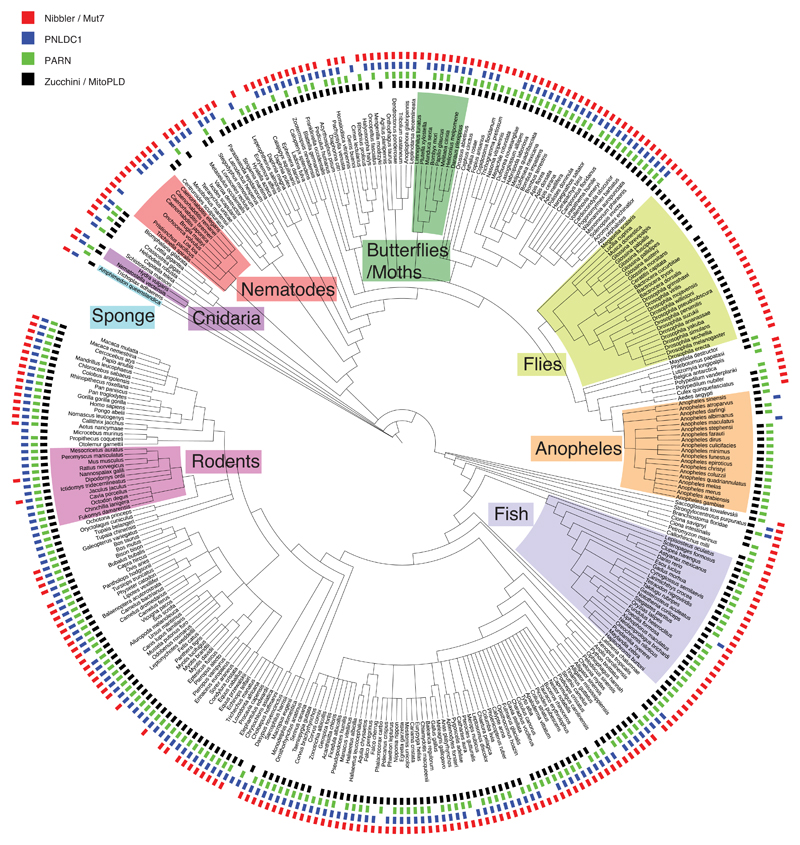
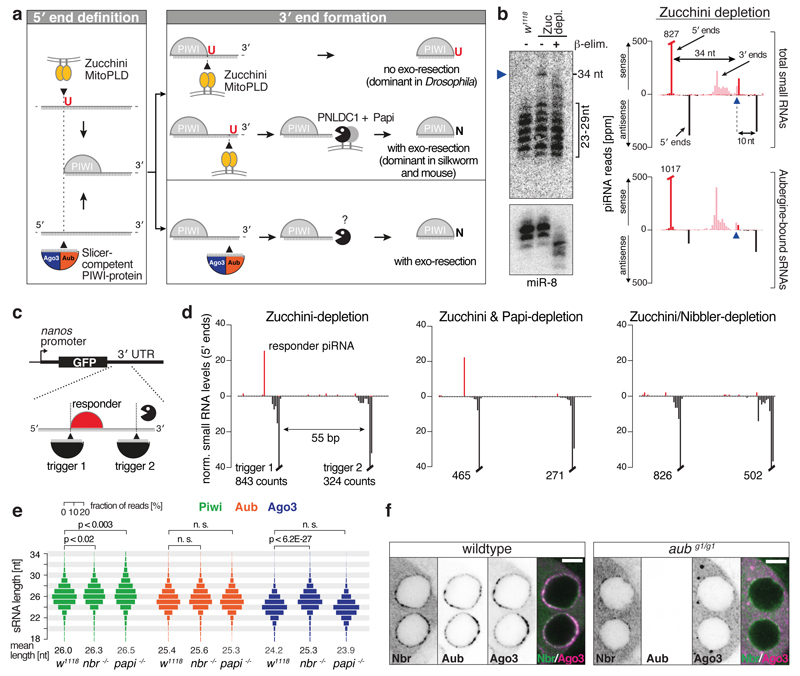
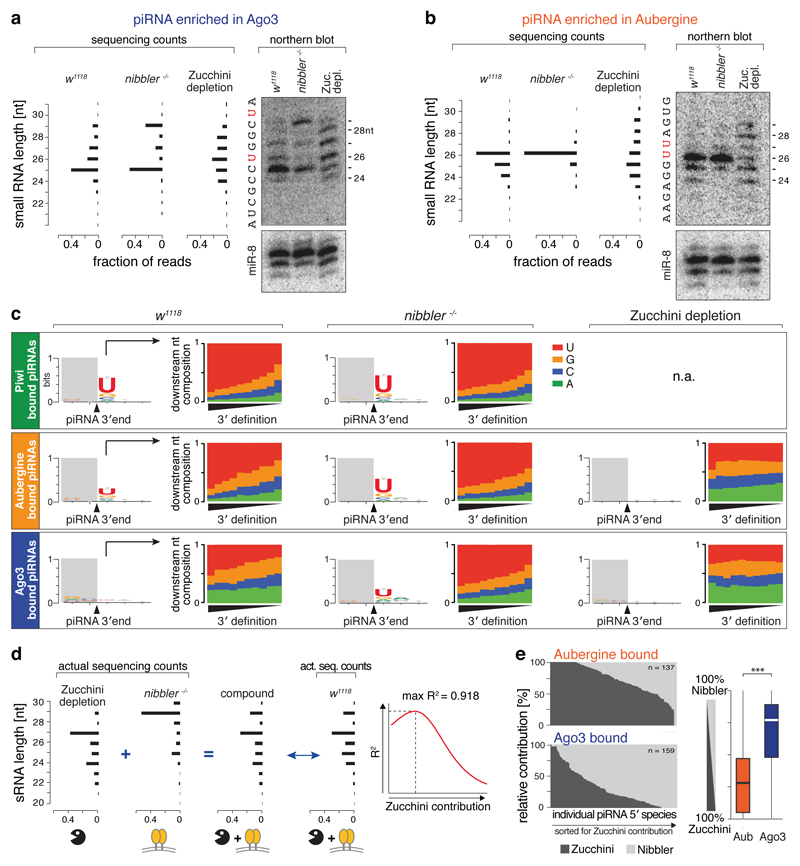
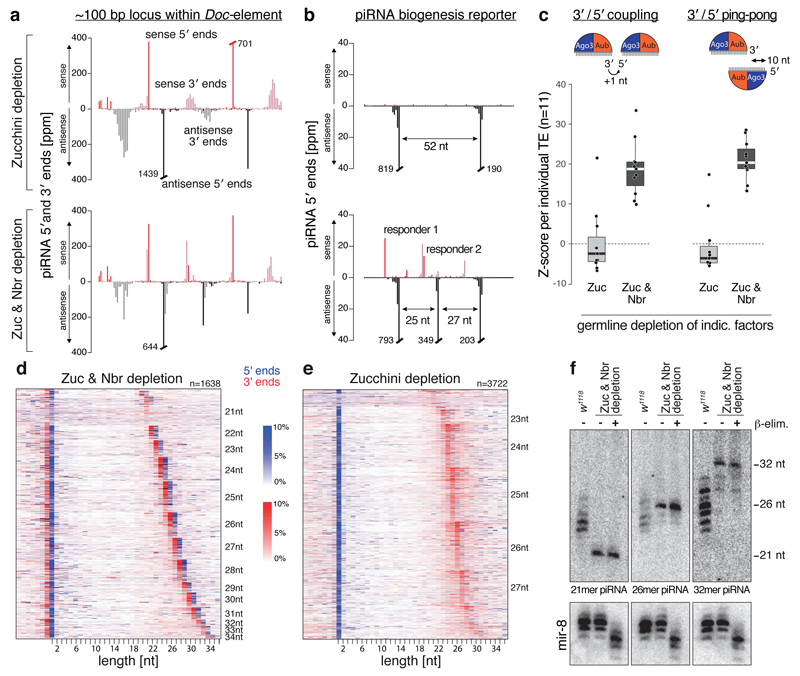
Small regulatory RNAs guide Argonaute (Ago) proteins in a sequence-specific manner to their targets and therefore have important roles in eukaryotic gene silencing. Of the three small RNA classes, microRNAs and short interfering RNAs are processed from double-stranded precursors into defined 21- to 23-mers by Dicer, an endoribonuclease with intrinsic ruler function. PIWI-interacting RNAs (piRNAs)-the 22-30-nt-long guides for PIWI-clade Ago proteins that silence transposons in animal gonads-are generated independently of Dicer from single-stranded precursors. piRNA 5' ends are defined either by Zucchini, the Drosophila homologue of mitoPLD-a mitochondria-anchored endonuclease, or by piRNA-guided target cleavage. Formation of piRNA 3' ends is poorly understood. Here we report that two genetically and mechanistically distinct pathways generate piRNA 3' ends in Drosophila. The initiating nucleases are either Zucchini or the PIWI-clade proteins Aubergine (Aub) or Ago3. While Zucchini-mediated cleavages directly define mature piRNA 3' ends, Aub/Ago3-mediated cleavages liberate pre-piRNAs that require extensive resection by the 3'-to-5' exoribonuclease Nibbler (Drosophila homologue of Mut-7). The relative activity of these two pathways dictates the extent to which piRNAs are directed to cytoplasmic or nuclear PIWI-clade proteins and thereby sets the balance between post-transcriptional and transcriptional silencing. Notably, loss of both Zucchini and Nibbler reveals a minimal, Argonaute-driven small RNA biogenesis pathway in which piRNA 5' and 3' ends are directly produced by closely spaced Aub/Ago3-mediated cleavage events. Our data reveal a coherent model for piRNA biogenesis, and should aid the mechanistic dissection of the processes that govern piRNA 3'-end formation.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
