

Reconstitution of Targeted Deadenylation by the Ccr4-Not Complex and the YTH Domain Protein Mmi1
- PMID: 27851962
- PMCID: PMC5120349
- DOI: 10.1016/j.celrep.2016.10.066
Reconstitution of Targeted Deadenylation by the Ccr4-Not Complex and the YTH Domain Protein Mmi1
Abstract
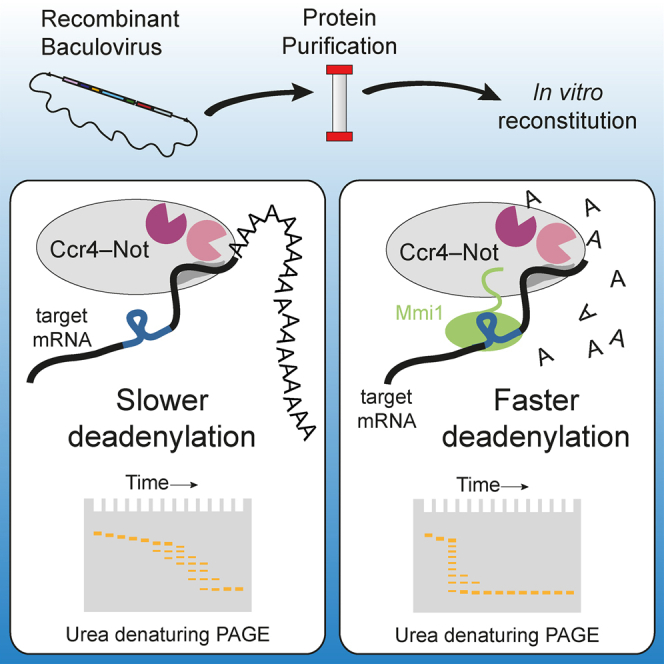
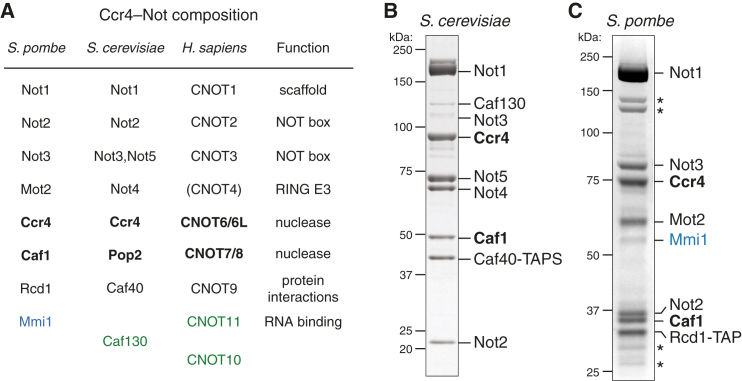
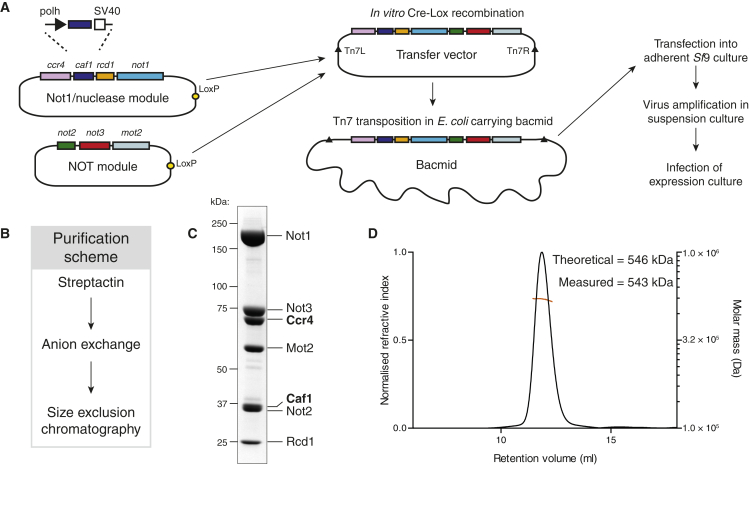
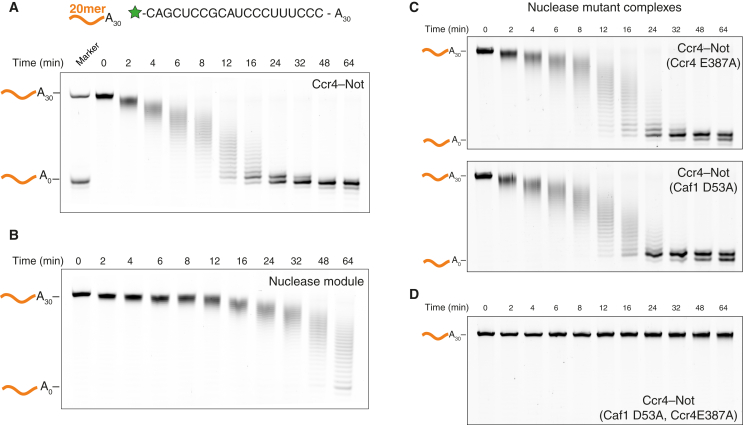
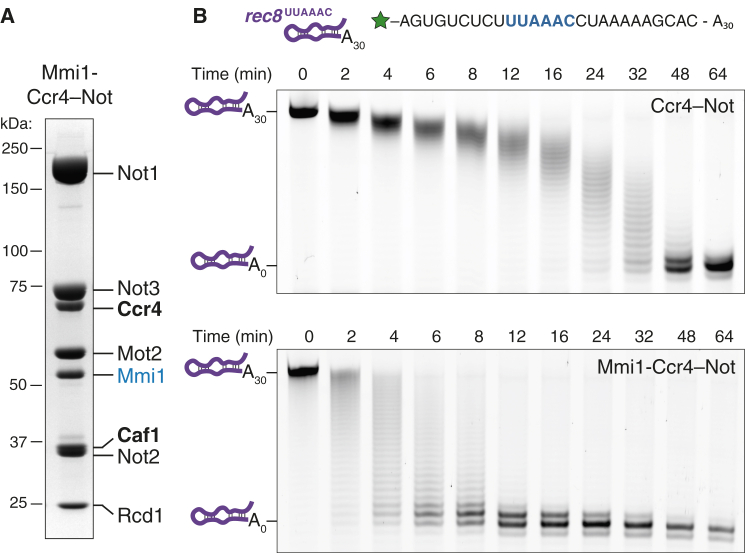
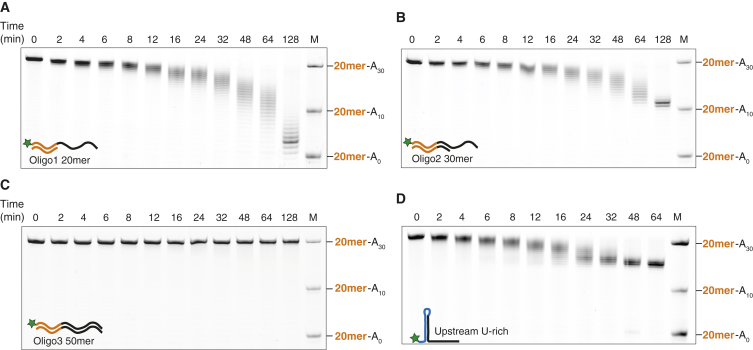
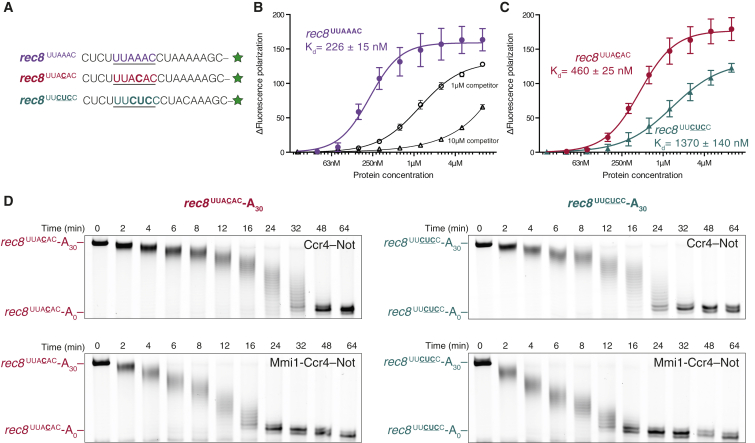
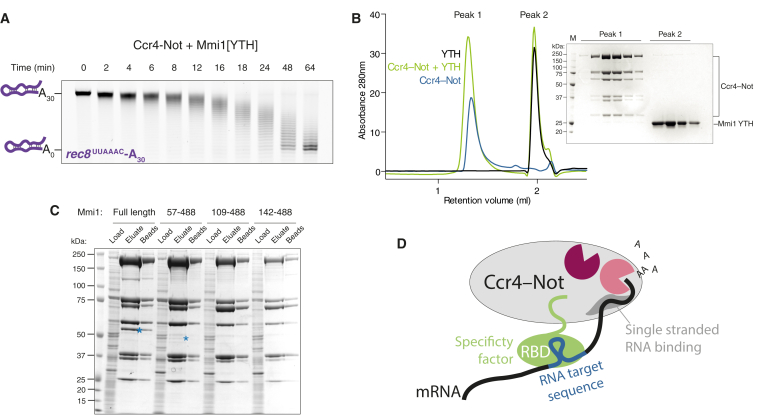
Ccr4-Not is a conserved protein complex that shortens the 3' poly(A) tails of eukaryotic mRNAs to regulate transcript stability and translation into proteins. RNA-binding proteins are thought to facilitate recruitment of Ccr4-Not to certain mRNAs, but lack of an in-vitro-reconstituted system has slowed progress in understanding the mechanistic details of this specificity. Here, we generate a fully recombinant Ccr4-Not complex that removes poly(A) tails from RNA substrates. The intact complex is more active than the exonucleases alone and has an intrinsic preference for certain RNAs. The RNA-binding protein Mmi1 is highly abundant in preparations of native Ccr4-Not. We demonstrate a high-affinity interaction between recombinant Ccr4-Not and Mmi1. Using in vitro assays, we show that Mmi1 accelerates deadenylation of target RNAs. Together, our results support a model whereby both RNA-binding proteins and the sequence context of mRNAs influence deadenylation rate to regulate gene expression.
Keywords: Ccr4-Not; RNA; exonuclease; gene expression; poly(A) tail.
Copyright © 2016 MRC Laboratory of Molecular Biology. Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
mRNA Deadenylation Is Coupled to Translation Rates by the Differential Activities of Ccr4-Not Nucleases.Mol Cell. 2018 Jun 21;70(6):1089-1100.e8. doi: 10.1016/j.molcel.2018.05.033. Mol Cell. 2018. PMID: 29932902 Free PMC article.
-
The intrinsic structure of poly(A) RNA determines the specificity of Pan2 and Caf1 deadenylases.Nat Struct Mol Biol. 2019 Jun;26(6):433-442. doi: 10.1038/s41594-019-0227-9. Epub 2019 May 20. Nat Struct Mol Biol. 2019. PMID: 31110294 Free PMC article.
-
RNA-binding proteins distinguish between similar sequence motifs to promote targeted deadenylation by Ccr4-Not.Elife. 2019 Jan 2;8:e40670. doi: 10.7554/eLife.40670. Elife. 2019. PMID: 30601114 Free PMC article.
-
Molecular Insights into mRNA Polyadenylation and Deadenylation.Int J Mol Sci. 2022 Sep 20;23(19):10985. doi: 10.3390/ijms231910985. Int J Mol Sci. 2022. PMID: 36232288 Free PMC article. Review.
-
The Ccr4-Not Complex: Architecture and Structural Insights.Subcell Biochem. 2017;83:349-379. doi: 10.1007/978-3-319-46503-6_13. Subcell Biochem. 2017. PMID: 28271483 Review.
Cited by
-
meiRNA, A Polyvalent Player in Fission Yeast Meiosis.Noncoding RNA. 2019 Sep 17;5(3):45. doi: 10.3390/ncrna5030045. Noncoding RNA. 2019. PMID: 31533287 Free PMC article. Review.
-
Regulation of the multisubunit CCR4-NOT deadenylase in the initiation of mRNA degradation.Curr Opin Struct Biol. 2022 Dec;77:102460. doi: 10.1016/j.sbi.2022.102460. Epub 2022 Sep 16. Curr Opin Struct Biol. 2022. PMID: 36116370 Free PMC article. Review.
-
The TUTase URT1 connects decapping activators and prevents the accumulation of excessively deadenylated mRNAs to avoid siRNA biogenesis.Nat Commun. 2021 Feb 26;12(1):1298. doi: 10.1038/s41467-021-21382-2. Nat Commun. 2021. PMID: 33637717 Free PMC article.
-
Disruption of the Mammalian Ccr4-Not Complex Contributes to Transcription-Mediated Genome Instability.Cells. 2023 Jul 17;12(14):1868. doi: 10.3390/cells12141868. Cells. 2023. PMID: 37508532 Free PMC article.
-
Structure and function of molecular machines involved in deadenylation-dependent 5'-3' mRNA degradation.Front Genet. 2023 Oct 9;14:1233842. doi: 10.3389/fgene.2023.1233842. eCollection 2023. Front Genet. 2023. PMID: 37876592 Free PMC article. Review.
References
-
- Basquin J., Roudko V.V., Rode M., Basquin C., Séraphin B., Conti E. Architecture of the nuclease module of the yeast Ccr4-not complex: the Not1-Caf1-Ccr4 interaction. Mol. Cell. 2012;48:207–218. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
