

Amyloid-β Peptide Nitrotyrosination Stabilizes Oligomers and Enhances NMDAR-Mediated Toxicity
- PMID: 27852777
- PMCID: PMC6705640
- DOI: 10.1523/JNEUROSCI.1081-16.2016
Amyloid-β Peptide Nitrotyrosination Stabilizes Oligomers and Enhances NMDAR-Mediated Toxicity
Abstract
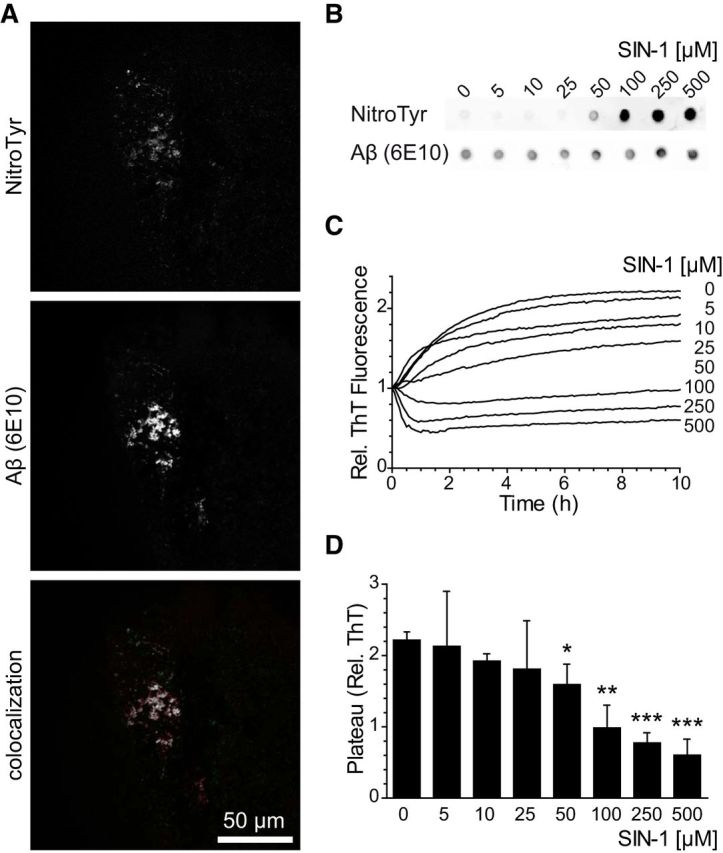
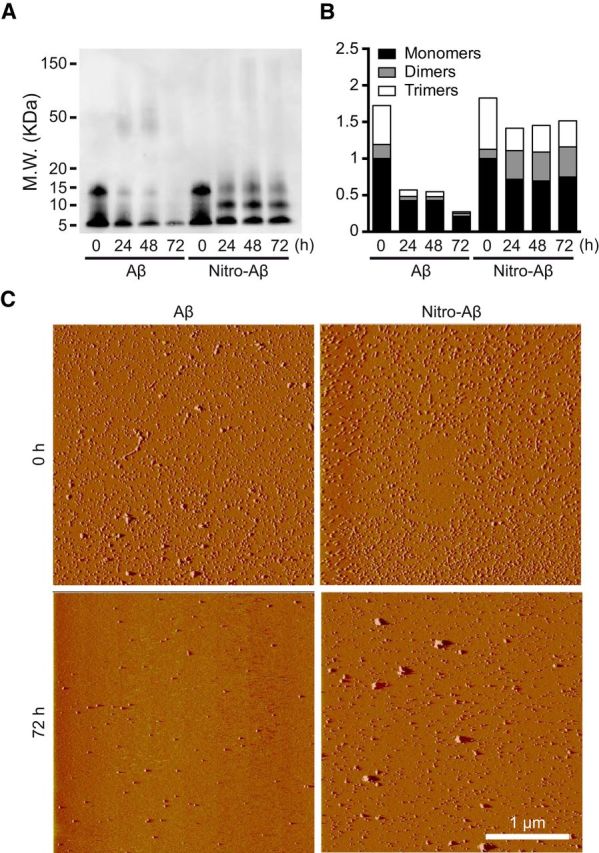
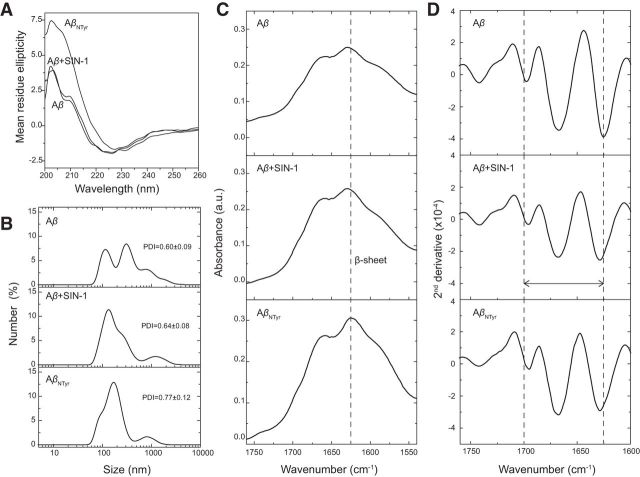
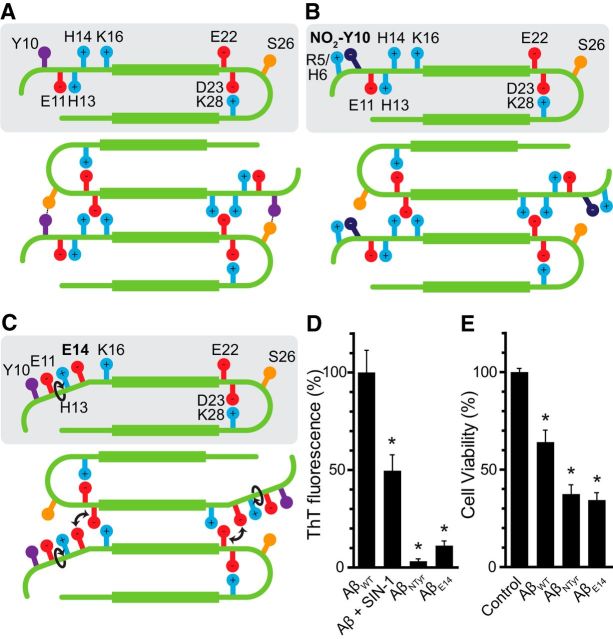
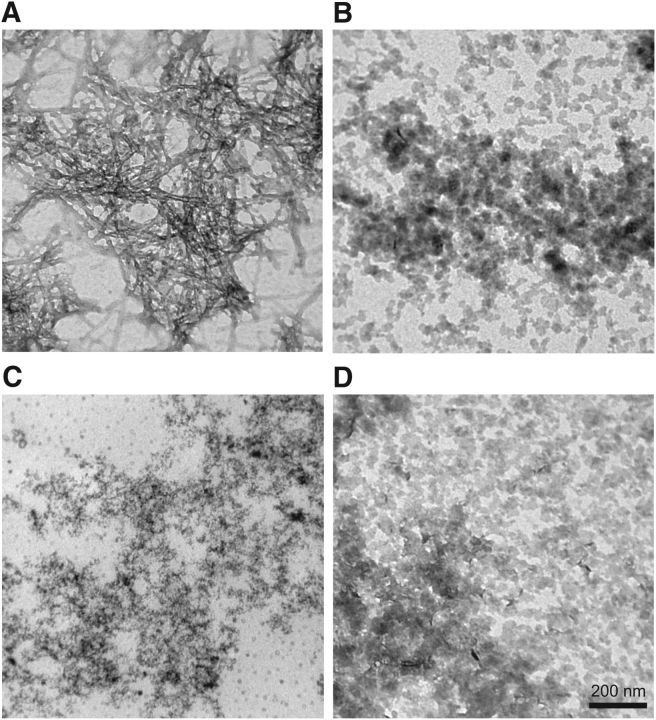
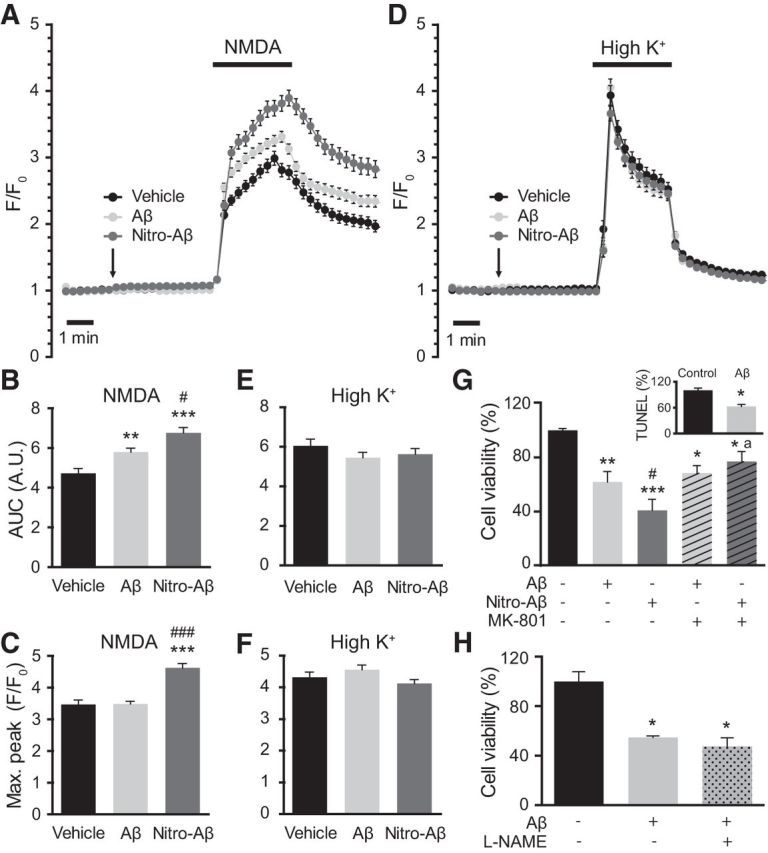
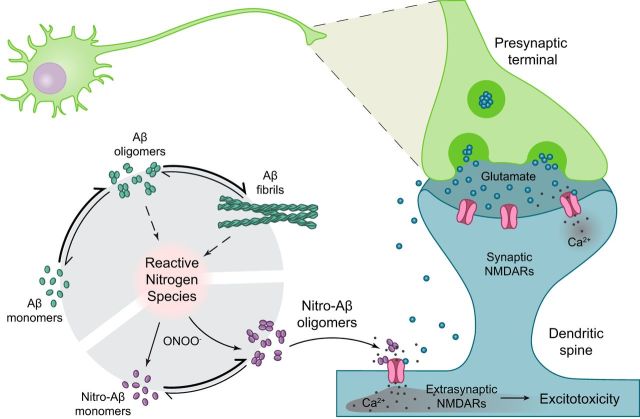
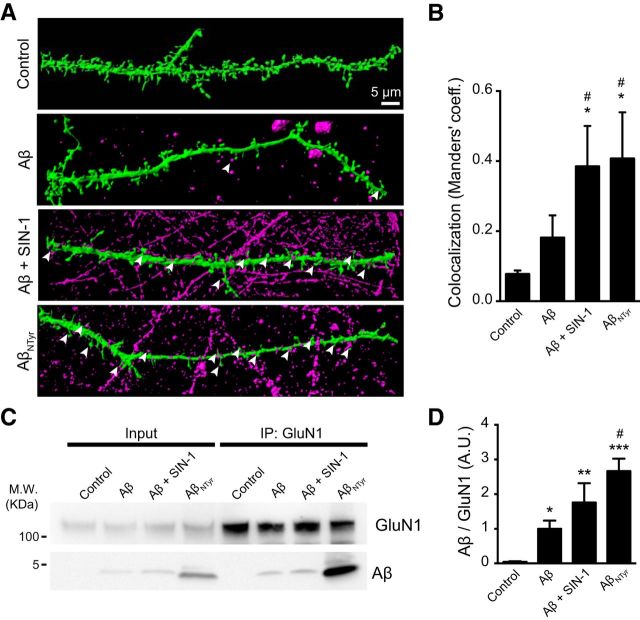
Alzheimer's disease (AD) is a neurodegenerative disorder characterized by the pathological aggregation of the amyloid-β peptide (Aβ). Monomeric soluble Aβ can switch from helicoidal to β-sheet conformation, promoting its assembly into oligomers and subsequently to amyloid fibrils. Oligomers are highly toxic to neurons and have been reported to induce synaptic transmission impairments. The progression from oligomers to fibrils forming senile plaques is currently considered a protective mechanism to avoid the presence of the highly toxic oligomers. Protein nitration is a frequent post-translational modification under AD nitrative stress conditions. Aβ can be nitrated at tyrosine 10 (Y10) by peroxynitrite. Based on our analysis of ThT binding, Western blot and electron and atomic force microscopy, we report that Aβ nitration stabilizes soluble, highly toxic oligomers and impairs the formation of fibrils. We propose a mechanism by which fibril elongation is interrupted upon Y10 nitration: Nitration disrupts fibril-forming folds by preventing H14-mediated bridging, as shown with an Aβ analog containing a single residue (H to E) replacement that mimics the behavior of nitrated Aβ related to fibril formation and neuronal toxicity. The pathophysiological role of our findings in AD was highlighted by the study of these nitrated oligomers on mouse hippocampal neurons, where an increased NMDAR-dependent toxicity of nitrated Aβ oligomers was observed. Our results show that Aβ nitrotyrosination is a post-translational modification that increases Aβ synaptotoxicity.
Significance statement: We report that nitration (i.e., the irreversible addition of a nitro group) of the Alzheimer-related peptide amyloid-β (Aβ) favors the stabilization of highly toxic oligomers and inhibits the formation of Aβ fibrils. The nitrated Aβ oligomers are more toxic to neurons due to increased cytosolic calcium levels throughout their action on NMDA receptors. Sustained elevated calcium levels trigger excitotoxicity, a characteristic event in Alzheimer's disease.
Keywords: Alzheimer; NMDA Rc; amyloid; nitrotyrosination; oligomers; peroxynitrite.
Copyright © 2016 the authors 0270-6474/16/3611693-11$15.00/0.
Figures
References
-
- Beckman JS, Koppenol WH. Nitric oxide, superoxide, and peroxynitrite: the good, the bad, and ugly. Am J Physiol. 1996;271:C1424–C1437. - PubMed
-
- Bitan G, Teplow DB. Preparation of aggregate-free, low molecular weight amyloid-beta for assembly and toxicity assays. Methods Mol Biol. 2005;299:3–9. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources