

Honey Bee Deformed Wing Virus Structures Reveal that Conformational Changes Accompany Genome Release
- PMID: 27852845
- PMCID: PMC5215330
- DOI: 10.1128/JVI.01795-16
Honey Bee Deformed Wing Virus Structures Reveal that Conformational Changes Accompany Genome Release
Abstract
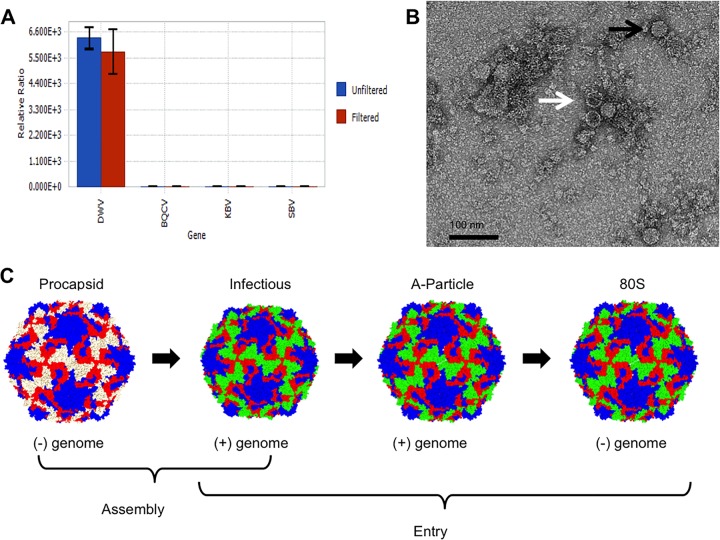
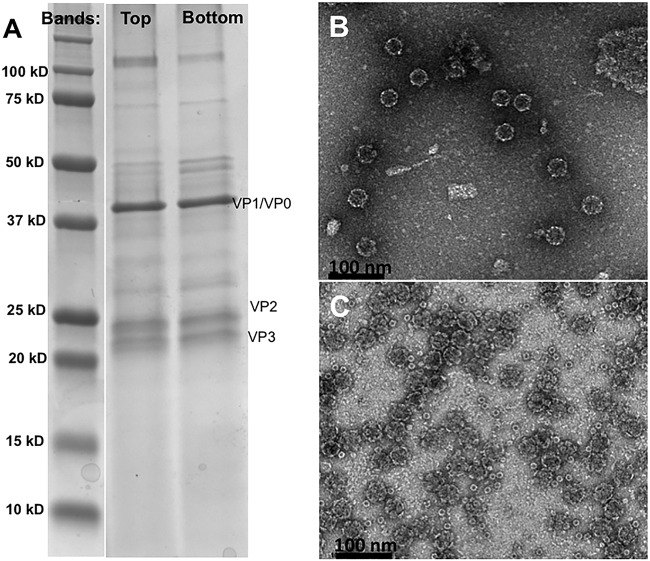
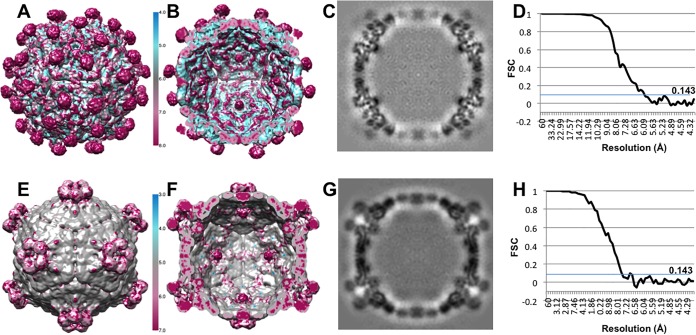
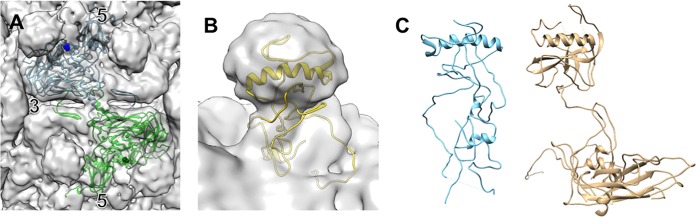
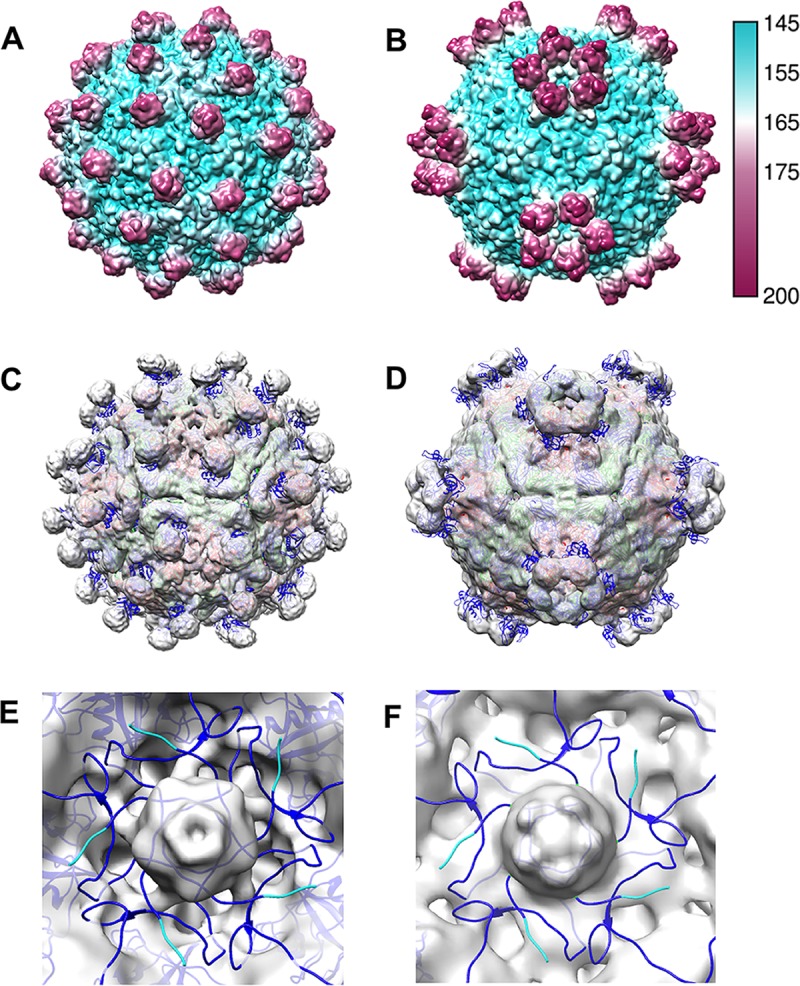
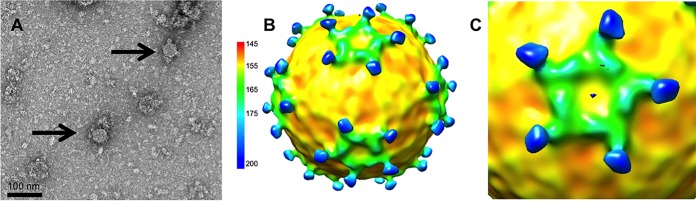
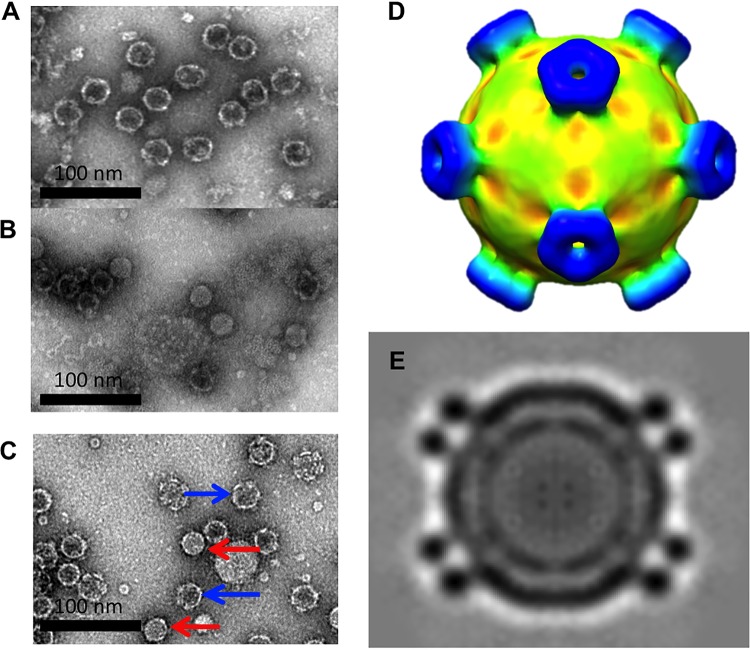
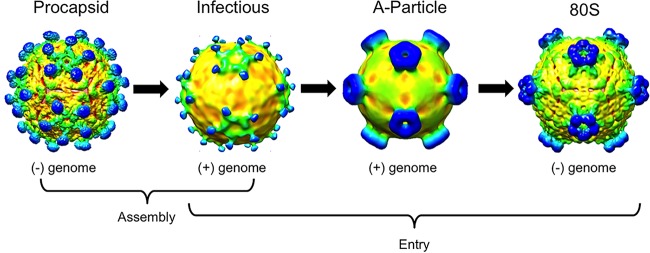
The picornavirus-like deformed wing virus (DWV) has been directly linked to colony collapse; however, little is known about the mechanisms of host attachment or entry for DWV or its molecular and structural details. Here we report the three-dimensional (3-D) structures of DWV capsids isolated from infected honey bees, including the immature procapsid, the genome-filled virion, the putative entry intermediate (A-particle), and the empty capsid that remains after genome release. The capsids are decorated by large spikes around the 5-fold vertices. The 5-fold spikes had an open flower-like conformation for the procapsid and genome-filled capsids, whereas the putative A-particle and empty capsids that had released the genome had a closed tube-like spike conformation. Between the two conformations, the spikes undergo a significant hinge-like movement that we predicted using a Robetta model of the structure comprising the spike. We conclude that the spike structures likely serve a function during host entry, changing conformation to release the genome, and that the genome may escape from a 5-fold vertex to initiate infection. Finally, the structures illustrate that, similarly to picornaviruses, DWV forms alternate particle conformations implicated in assembly, host attachment, and RNA release.
Importance: Honey bees are critical for global agriculture, but dramatic losses of entire hives have been reported in numerous countries since 2006. Deformed wing virus (DWV) and infestation with the ectoparasitic mite Varroa destructor have been linked to colony collapse disorder. DWV was purified from infected adult worker bees to pursue biochemical and structural studies that allowed the first glimpse into the conformational changes that may be required during transmission and genome release for DWV.
Keywords: 5-fold spikes; 80S; DWV; conformation change; deformed wing virus; honey bee; insect; life cycle; picornavirus; procapsid.
Copyright © 2017 American Society for Microbiology.
Figures



References
-
- Mondet F, de Miranda JR, Kretzschmar A, Le Conte Y, Mercer AR. 2014. On the front line: quantitative virus dynamics in honeybee (Apis mellifera L.) colonies along a new expansion front of the parasite Varroa destructor. PLoS Pathog 10:e1004323. doi: 10.1371/journal.ppat.1004323. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
