

A Novel Rabies Vaccine Expressing CXCL13 Enhances Humoral Immunity by Recruiting both T Follicular Helper and Germinal Center B Cells
- PMID: 27852854
- PMCID: PMC5244327
- DOI: 10.1128/JVI.01956-16
A Novel Rabies Vaccine Expressing CXCL13 Enhances Humoral Immunity by Recruiting both T Follicular Helper and Germinal Center B Cells
Abstract
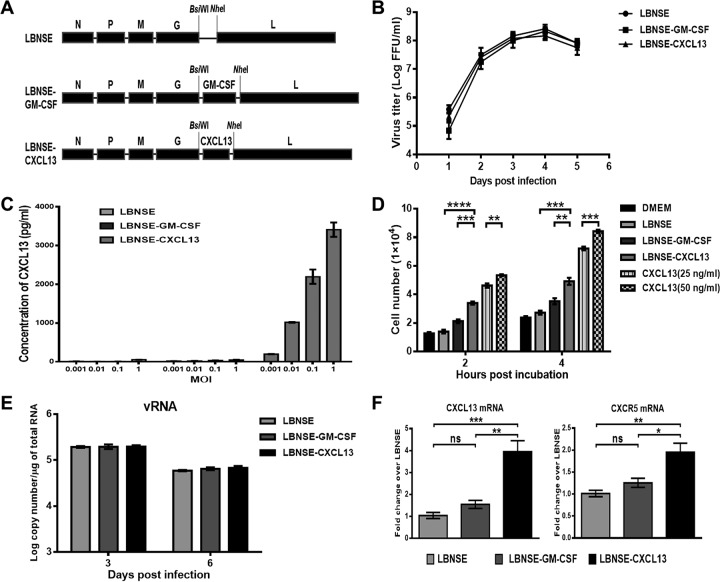
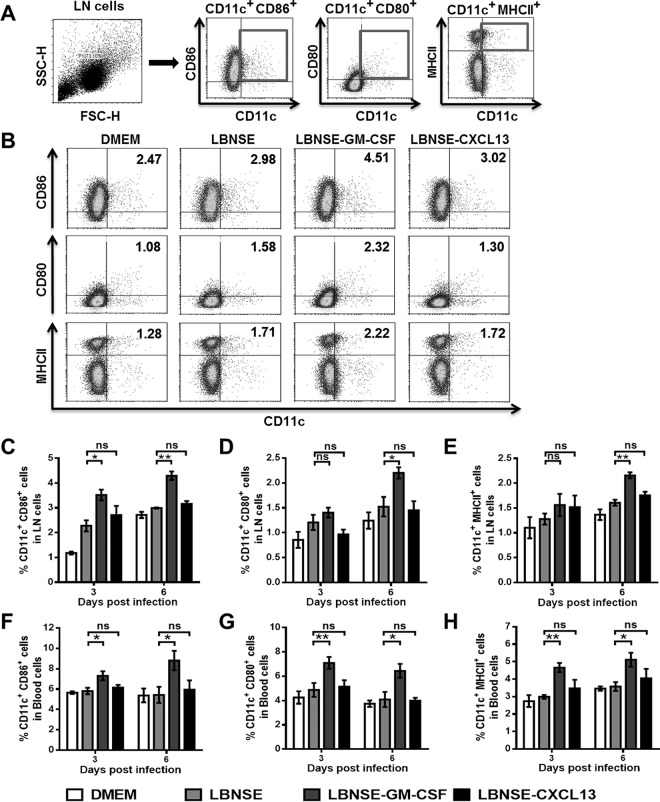
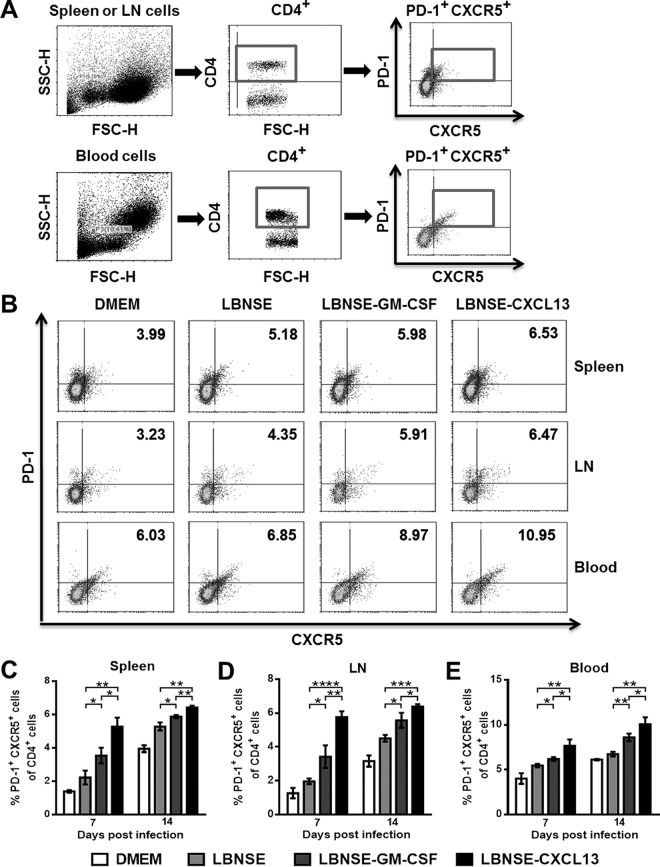
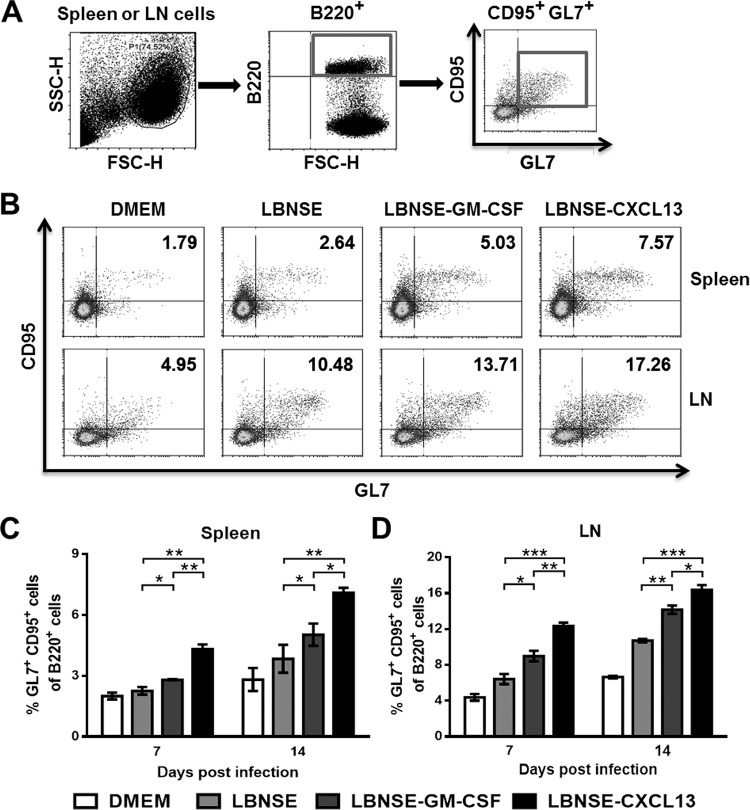
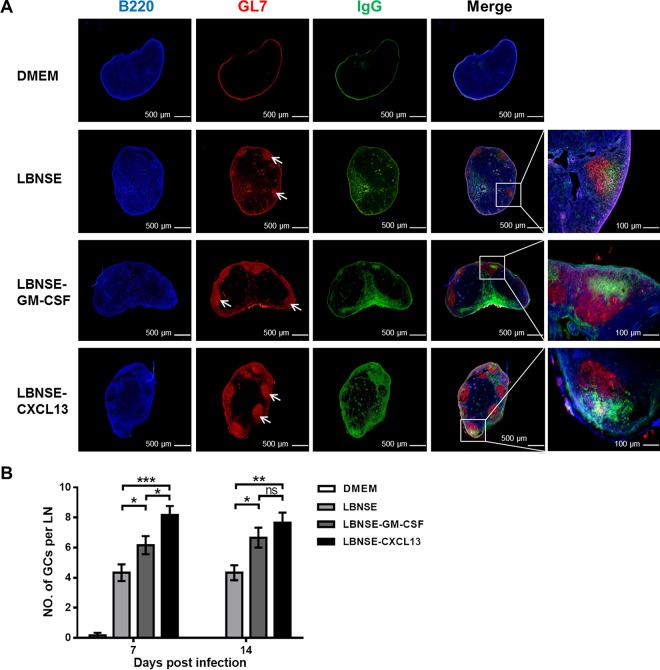
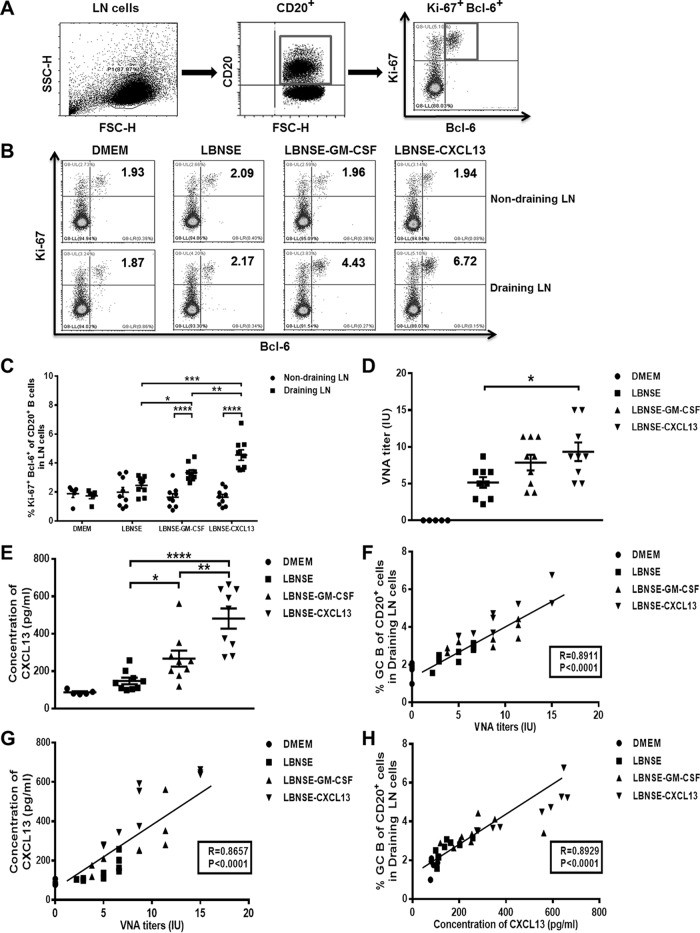
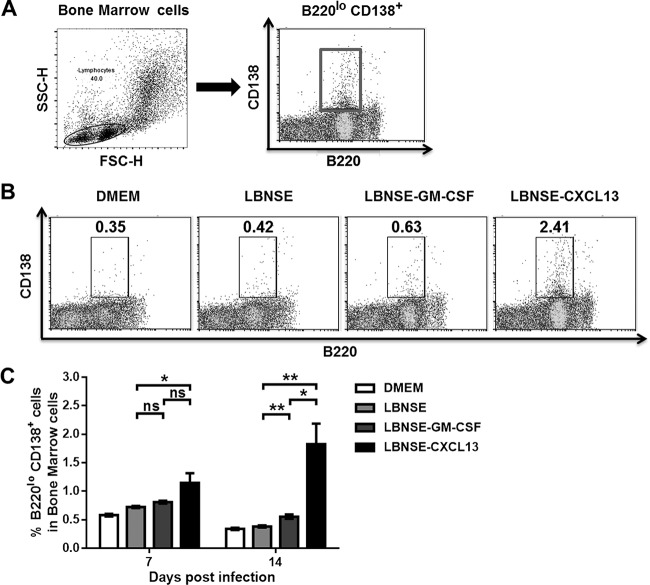
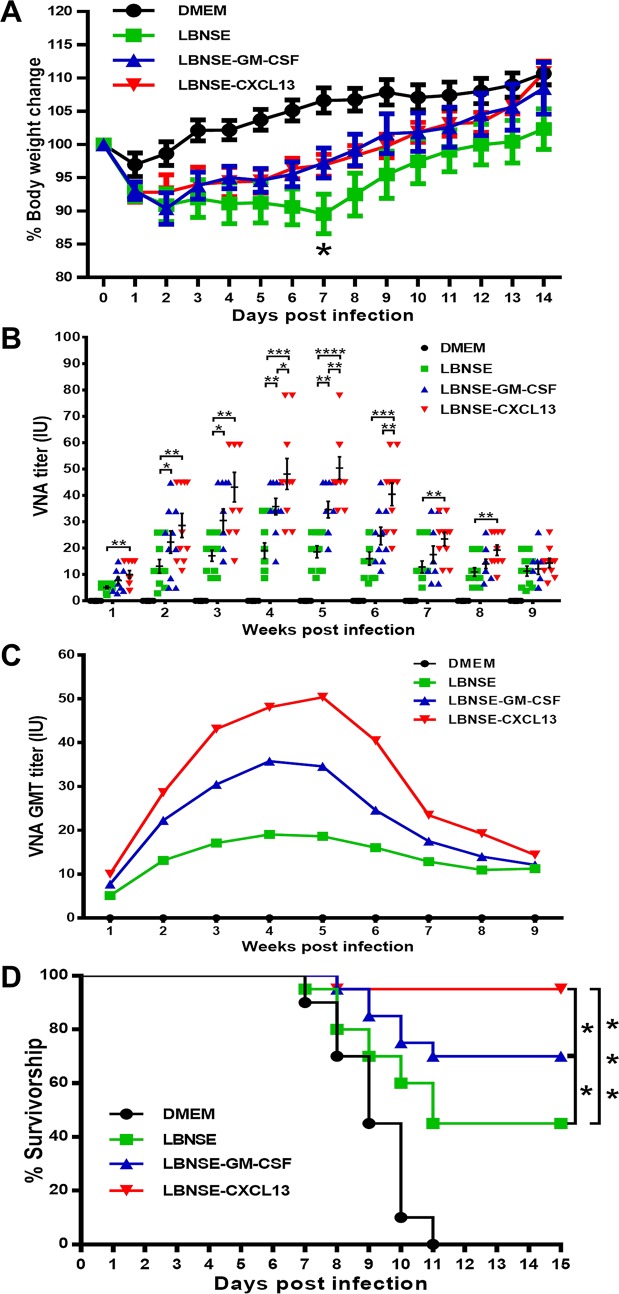
Rabies remains a public health threat in most parts of the world, and approximately 99% of the cases are transmitted by dogs. There is an urgent need to develop an efficacious and affordable vaccine to control canine-transmitted rabies in developing countries. Our previous studies demonstrate that overexpression of chemokines/cytokines such as CCL-3 (MIP-1α) and granulocyte-macrophage colony-stimulating factor (GM-CSF) can enhance the immunogenicity of rabies vaccines. In the present study, the chemokine CXCL13 was inserted into the genome of the recombinant rabies virus (rRABV) strain LBNSE, and the effect of the chemokine CXCL13 on the immunogenicity of RABV was investigated. It was found that LBNSE-CXCL13 recruited follicular helper T (Tfh) and germinal center (GC) B cells, promoted the formation of GCs, and increased the population of plasma cells in immunized mice. Further studies showed that mice immunized with LBNSE-CXCL13 produced more rabies virus-neutralizing antibodies (VNAs) and developed better protection than those immunized with the parent virus LBNSE or the GM-CSF-expressing RABV (LBNSE-GM-CSF). Collectively, these findings provide a better understanding of the role of CXCL13 expression in the immunogenicity of the RABV, which may help in designing more-efficacious rabies vaccines.
Importance: Rabies is endemic in most parts of the world, and more effort is needed to develop affordable and effective vaccines to control or eliminate this disease. The chemokine CXCL13 recruits both Tfh and B cells, which is essential for the homing of Tfh cells and the development of B cell follicles. In this study, the effect of the overexpression of CXCL13 on the immunogenicity of the RABV was evaluated in a mouse model. We found that CXCL13 expression promoted humoral immunity by recruiting Tfh and GC B cells, facilitating the formation of GCs, and increasing the number of plasma cells. As expected, the overexpression of CXCL13 resulted in enhanced virus-neutralizing antibody (VNA) production and protection against a virulent RABV challenge. These findings provide a better understanding of the role of CXCL13 in RABV-induced immune responses, which will help in designing more efficacious rabies vaccines.
Keywords: CXCL13; T follicular helper cells; germinal center B cells; humoral immunity; rabies virus.
Copyright © 2017 American Society for Microbiology.
Figures
Similar articles
-
A Recombinant Rabies Virus Expressing Fms-like Tyrosine Kinase 3 Ligand (Flt3L) Induces Enhanced Immunogenicity in Mice.Virol Sin. 2019 Dec;34(6):662-672. doi: 10.1007/s12250-019-00144-x. Epub 2019 Jun 28. Virol Sin. 2019. PMID: 31254272 Free PMC article.
-
A novel oral rabies vaccine enhances the immunogenicity through increasing dendritic cells activation and germinal center formation by expressing U-OMP19 in a mouse model.Emerg Microbes Infect. 2021 Dec;10(1):913-928. doi: 10.1080/22221751.2021.1923341. Emerg Microbes Infect. 2021. PMID: 33905300 Free PMC article.
-
Recombinant rabies virus expressing IL-21 enhances immunogenicity through activation of T follicular helper cells and germinal centre B cells.J Gen Virol. 2016 Dec;97(12):3154-3160. doi: 10.1099/jgv.0.000646. Epub 2016 Oct 25. J Gen Virol. 2016. PMID: 27902338
-
T follicular helper cell heterogeneity: Time, space, and function.Immunol Rev. 2019 Mar;288(1):85-96. doi: 10.1111/imr.12740. Immunol Rev. 2019. PMID: 30874350 Free PMC article. Review.
-
With a Little Help from T Follicular Helper Friends: Humoral Immunity to Influenza Vaccination.J Immunol. 2019 Jan 15;202(2):360-367. doi: 10.4049/jimmunol.1800986. J Immunol. 2019. PMID: 30617117 Review.
Cited by
-
CXCL13 promotes broad immune responses induced by circular RNA vaccines.Proc Natl Acad Sci U S A. 2024 Oct 29;121(44):e2406434121. doi: 10.1073/pnas.2406434121. Epub 2024 Oct 22. Proc Natl Acad Sci U S A. 2024. PMID: 39436660 Free PMC article.
-
Modelling Lyssavirus Infections in Human Stem Cell-Derived Neural Cultures.Viruses. 2020 Mar 25;12(4):359. doi: 10.3390/v12040359. Viruses. 2020. PMID: 32218146 Free PMC article.
-
Virus-Like Vesicles Based on Semliki Forest Virus-Containing Rabies Virus Glycoprotein Make a Safe and Efficacious Rabies Vaccine Candidate in a Mouse Model.J Virol. 2021 Sep 27;95(20):e0079021. doi: 10.1128/JVI.00790-21. Epub 2021 Aug 4. J Virol. 2021. PMID: 34346765 Free PMC article.
-
Recombinant rabies virus expressing IL-15 enhances immunogenicity through promoting the activation of dendritic cells in mice.Virol Sin. 2017 Aug;32(4):317-327. doi: 10.1007/s12250-017-4036-1. Epub 2017 Aug 29. Virol Sin. 2017. PMID: 28861771 Free PMC article.
-
A single immunization with core-shell structured lipopolyplex mRNA vaccine against rabies induces potent humoral immunity in mice and dogs.Emerg Microbes Infect. 2023 Dec;12(2):2270081. doi: 10.1080/22221751.2023.2270081. Epub 2023 Nov 22. Emerg Microbes Infect. 2023. PMID: 37819147 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
