

Potential for Low-Pathogenic Avian H7 Influenza A Viruses To Replicate and Cause Disease in a Mammalian Model
- PMID: 27852855
- PMCID: PMC5244340
- DOI: 10.1128/JVI.01934-16
Potential for Low-Pathogenic Avian H7 Influenza A Viruses To Replicate and Cause Disease in a Mammalian Model
Abstract
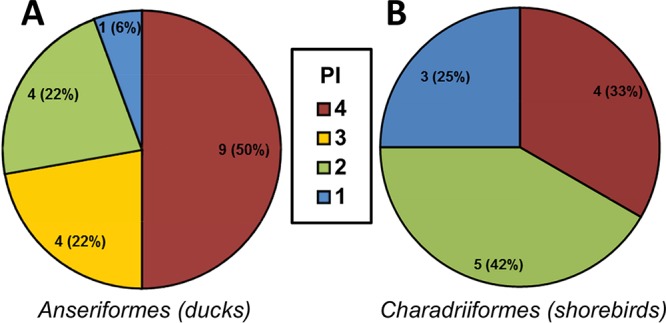
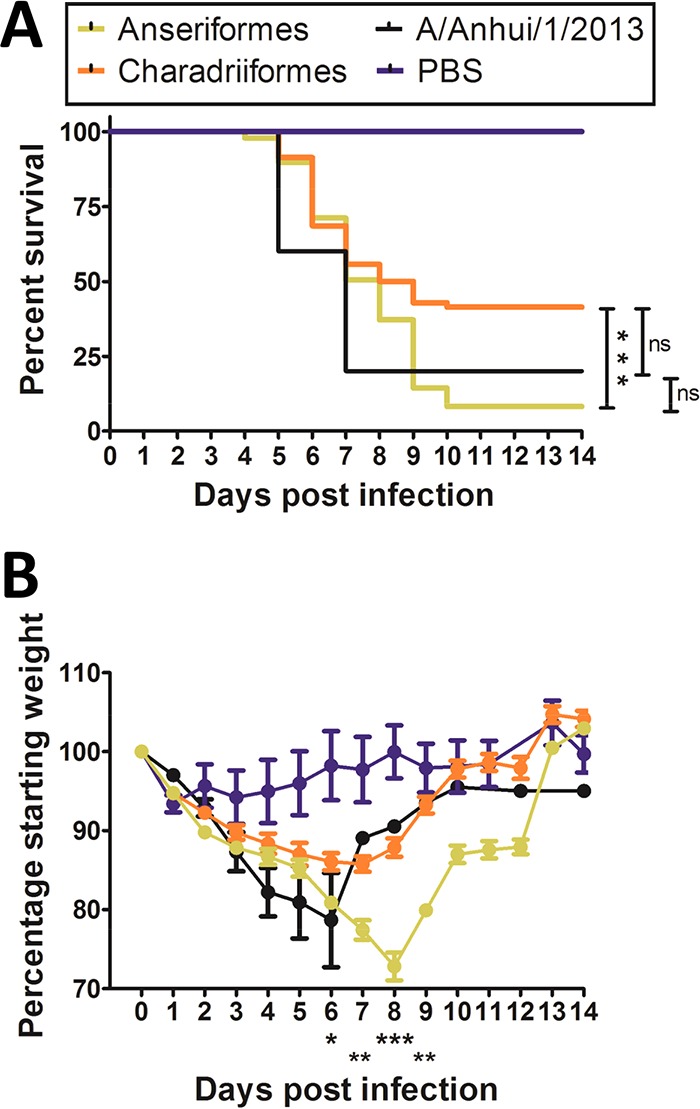
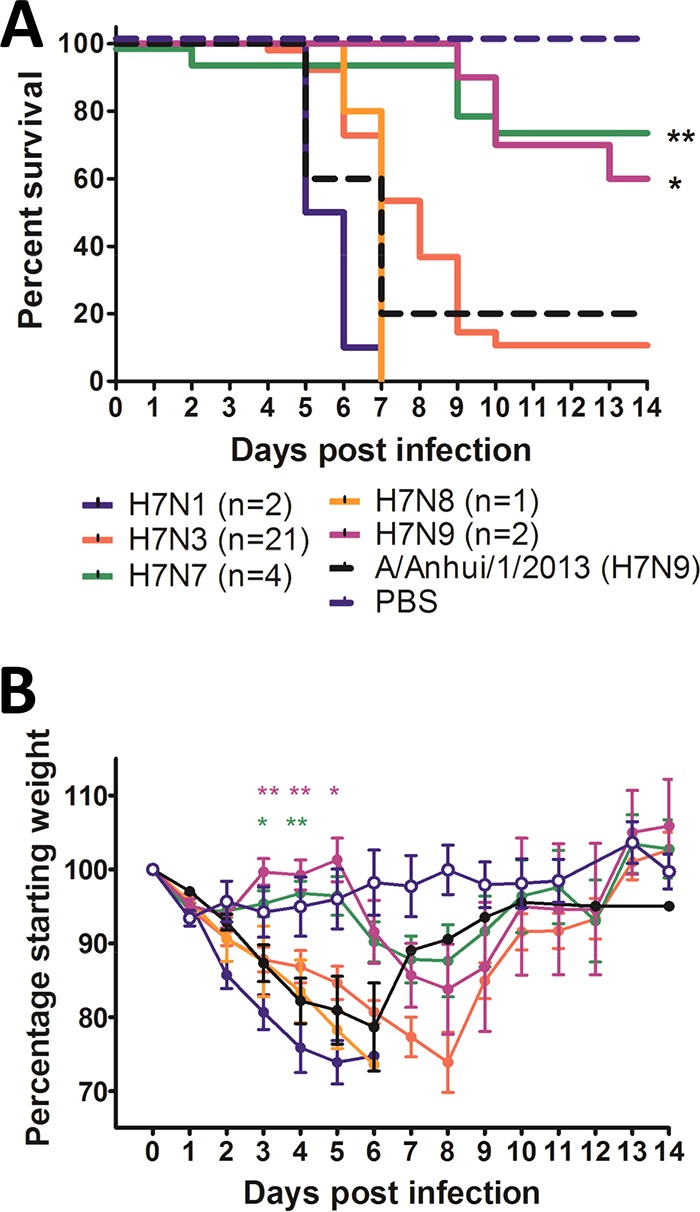
H7 subtype influenza A viruses are widely distributed and have been responsible for human infections and numerous outbreaks in poultry with significant impact. Despite this, the disease-causing potential of the precursor low-pathogenic (LP) H7 viruses from the wild bird reservoir has not been investigated. Our objective was to assess the disease-causing potential of 30 LP H7 viruses isolated from wild avian species in the United States and Canada using the DBA/2J mouse model. Without prior mammalian adaptation, the majority of viruses, 27 (90%), caused mortality in mice. Of these, 17 (56.7%) caused 100% mortality and 24 were of pathogenicity similar to that of A/Anhui/1/2013 (H7N9), which is highly pathogenic in mice. Viruses of duck origin were more pathogenic than those of shorebird origin, as 13 of 18 (72.2%) duck origin viruses caused 100% mortality while 4 of 12 (33.3%) shorebird origin viruses caused 100% mortality, despite there being no difference in mean lung viral titers between the groups. Replication beyond the respiratory tract was also evident, particularly in the heart and brain. Of the 16 viruses studied for fecal shedding, 11 were detected in fecal samples. These viruses exhibited a strong preference for avian-type α2,3-linked sialic acids; however, binding to mammalian-type α2,6-linked sialic acids was also detected. These findings indicate that LP avian H7 influenza A viruses are able to infect and cause disease in mammals without prior adaptation and therefore pose a potential public health risk.
Importance: Low-pathogenic (LP) avian H7 influenza A viruses are widely distributed in the avian reservoir and are the precursors of numerous outbreaks of highly pathogenic avian influenza viruses in commercial poultry farms. However, unlike highly pathogenic H7 viruses, the disease-causing potential of LP H7 viruses from the wild bird reservoir has not been investigated. To address this, we studied 30 LP avian H7 viruses isolated from wild avian species in the United States and Canada using the DBA/2J mouse model. Surprisingly, the majority of these viruses, 90%, caused mortality in mice without prior mammalian adaptation, and 56.7% caused 100% mortality. There was also evidence of spread beyond the respiratory tract and fecal shedding. Therefore, the disease-causing potential of LP avian H7 influenza A viruses in mammals may be underestimated, and these viruses therefore pose a potential public health risk.
Keywords: H7; avian viruses; influenza; viral pathogenesis.
Copyright © 2017 American Society for Microbiology.
Figures
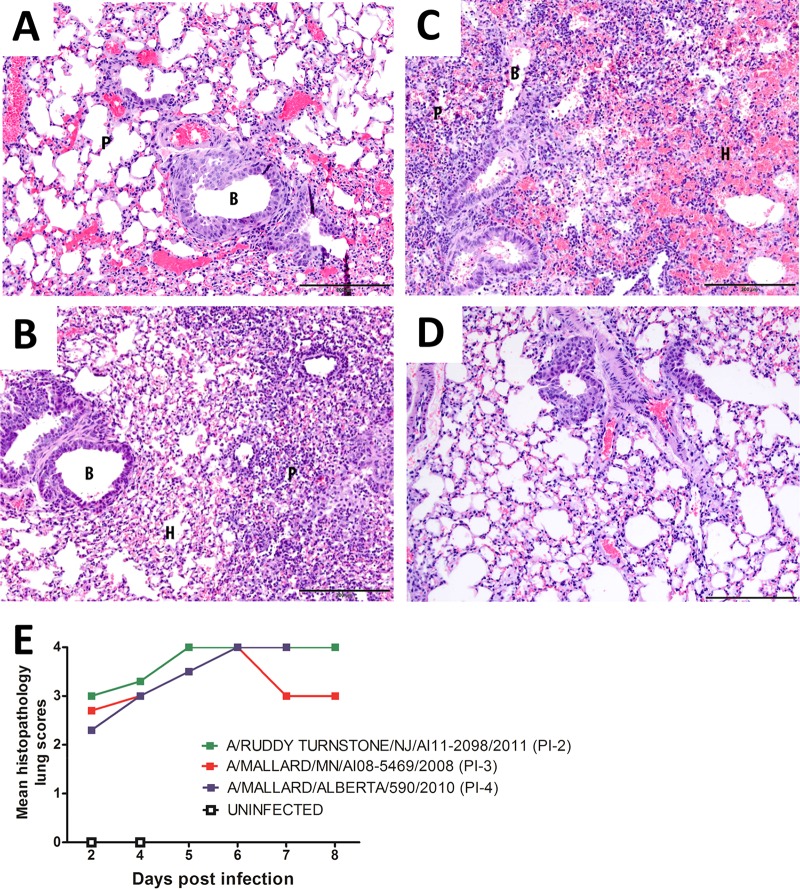
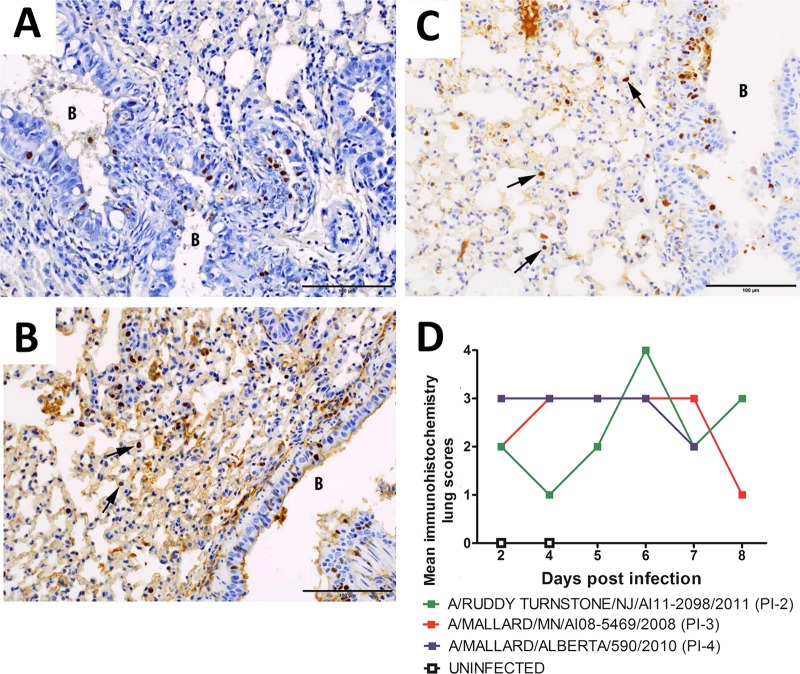

References
-
- Freidl GS, Meijer A, de Bruin E, de Nardi M, Munoz O, Capua I, Breed AC, Harris K, Hill A, Kosmider R, Banks J, von Dobschuetz S, Stark K, Wieland B, Stevens K, van der Werf S, Enouf V, van der Meulen K, Van Reeth K, Dauphin G, Koopmans M, FLURISK Consortium. 2014. Influenza at the animal-human interface: a review of the literature for virological evidence of human infection with swine or avian influenza viruses other than A(H5N1). Euro Surveill 19:20793. doi: 10.2807/1560-7917.ES2014.19.18.20793. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
