

Nuclear Export Signal Masking Regulates HIV-1 Rev Trafficking and Viral RNA Nuclear Export
- PMID: 27852860
- PMCID: PMC5244332
- DOI: 10.1128/JVI.02107-16
Nuclear Export Signal Masking Regulates HIV-1 Rev Trafficking and Viral RNA Nuclear Export
Abstract
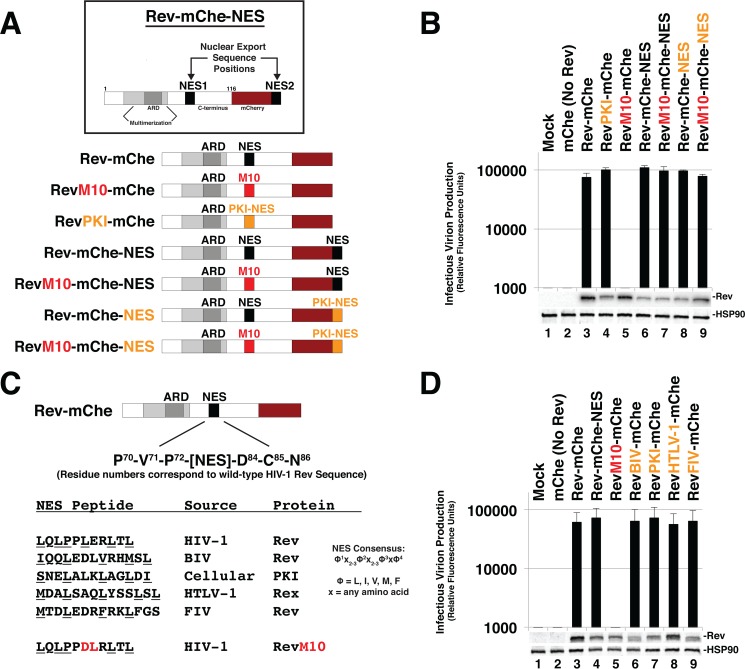
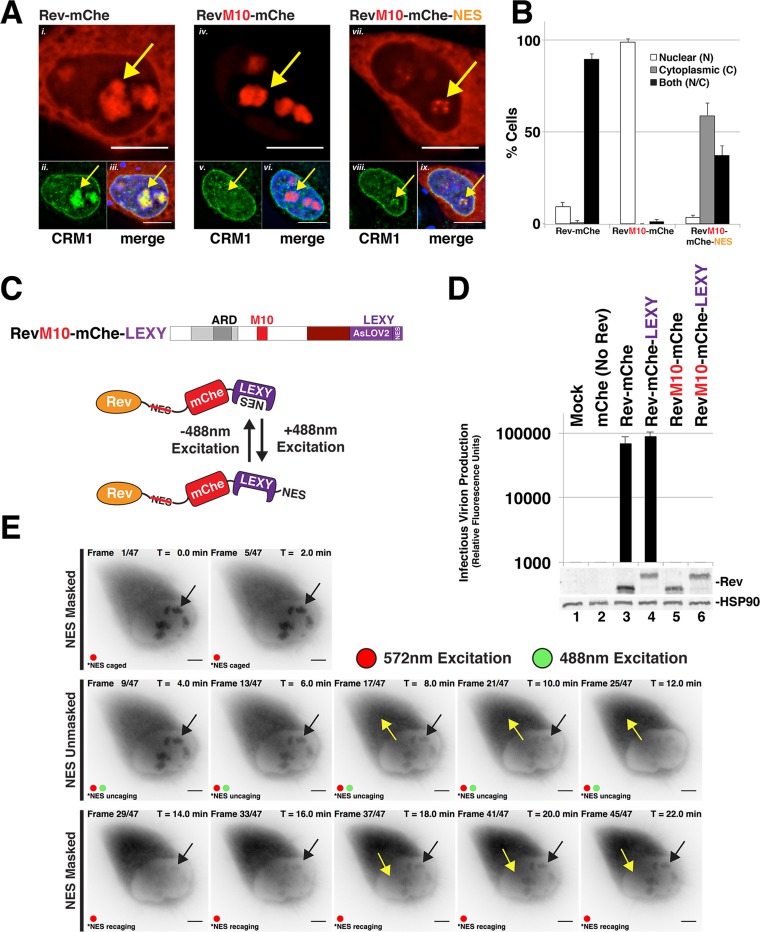
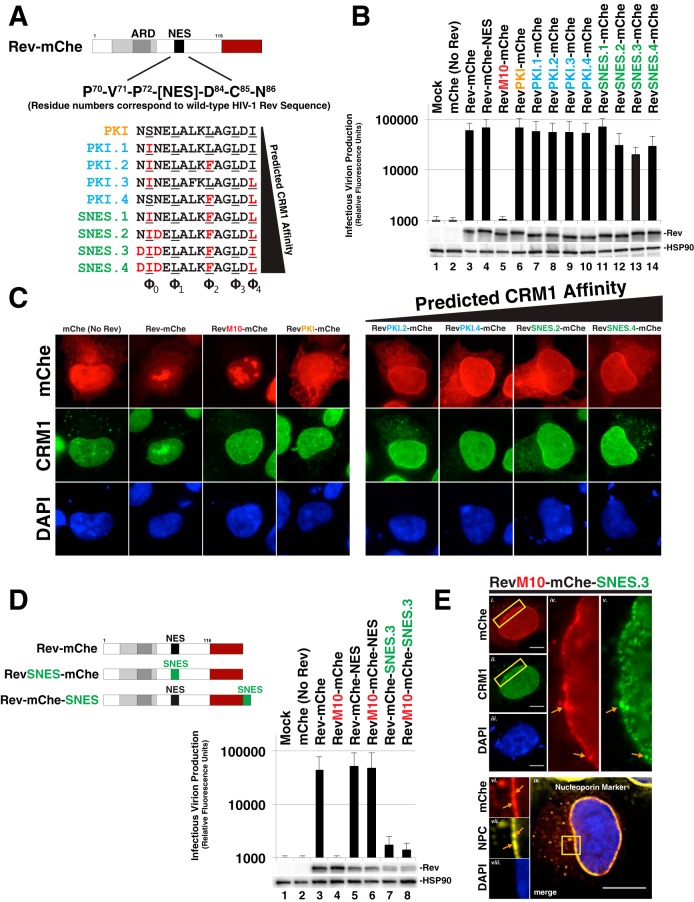
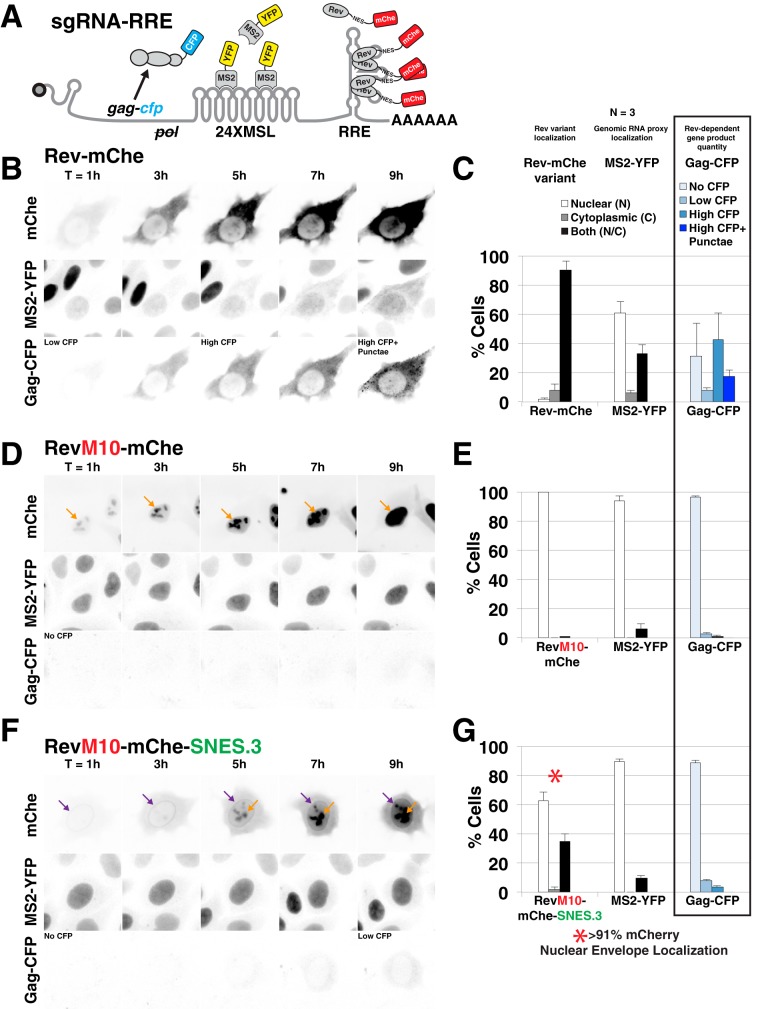
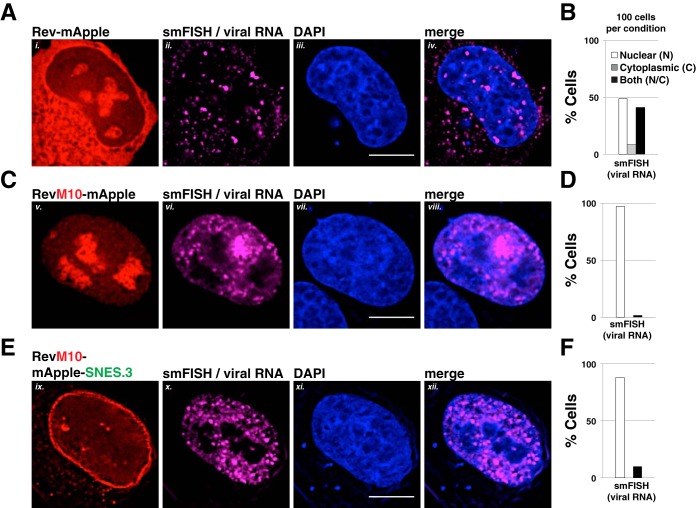
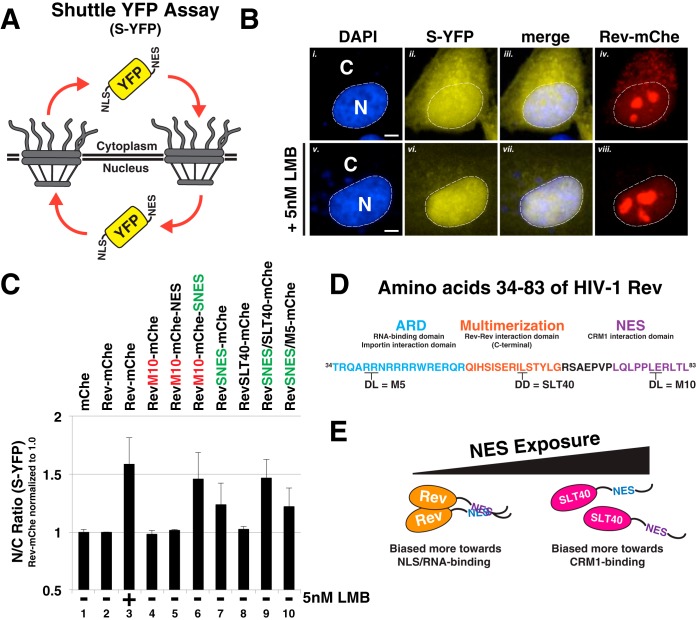
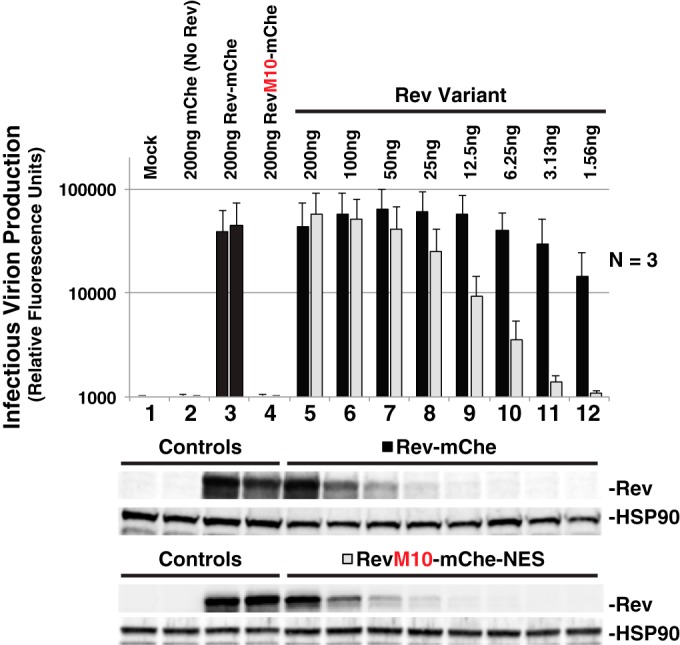
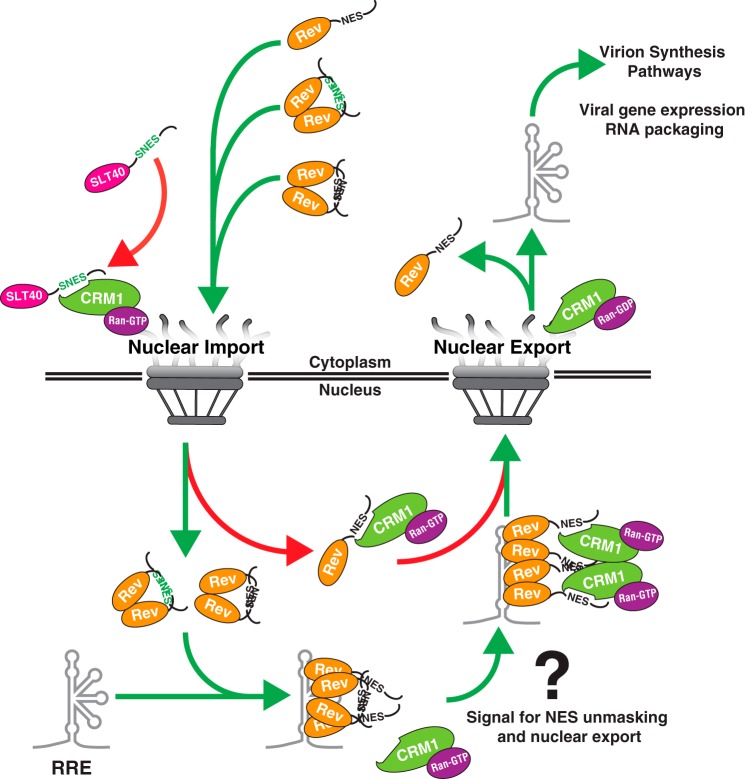
HIV-1's Rev protein forms a homo-oligomeric adaptor complex linking viral RNAs to the cellular CRM1/Ran-GTP nuclear export machinery through the activity of Rev's prototypical leucine-rich nuclear export signal (NES). In this study, we used a functional fluorescently tagged Rev fusion protein as a platform to study the effects of modulating Rev NES identity, number, position, or strength on Rev subcellular trafficking, viral RNA nuclear export, and infectious virion production. We found that Rev activity was remarkably tolerant of diverse NES sequences, including supraphysiological NES (SNES) peptides that otherwise arrest CRM1 transport complexes at nuclear pores. Rev's ability to tolerate a SNES was both position and multimerization dependent, an observation consistent with a model wherein Rev self-association acts to transiently mask the NES peptide(s), thereby biasing Rev's trafficking into the nucleus. Combined imaging and functional assays also indicated that NES masking underpins Rev's well-known tendency to accumulate at the nucleolus, as well as Rev's capacity to activate optimal levels of late viral gene expression. We propose that Rev multimerization and NES masking regulates Rev's trafficking to and retention within the nucleus even prior to RNA binding.
Importance: HIV-1 infects more than 34 million people worldwide causing >1 million deaths per year. Infectious virion production is activated by the essential viral Rev protein that mediates nuclear export of intron-bearing late-stage viral mRNAs. Rev's shuttling into and out of the nucleus is regulated by the antagonistic activities of both a peptide-encoded N-terminal nuclear localization signal and C-terminal nuclear export signal (NES). How Rev and related viral proteins balance strong import and export activities in order to achieve optimal levels of viral gene expression is incompletely understood. We provide evidence that multimerization provides a mechanism by which Rev transiently masks its NES peptide, thereby biasing its trafficking to and retention within the nucleus. Targeted pharmacological disruption of Rev-Rev interactions should perturb multiple Rev activities, both Rev-RNA binding and Rev's trafficking to the nucleus in the first place.
Keywords: CRM1; Gag; RNA trafficking; Rev; exportin-1; human immunodeficiency virus; nuclear export signal; nuclear pore complex; nucleolus; retroviruses.
Copyright © 2017 American Society for Microbiology.
Figures
Similar articles
-
Cooperativity among Rev-associated nuclear export signals regulates HIV-1 gene expression and is a determinant of virus species tropism.J Virol. 2014 Dec;88(24):14207-21. doi: 10.1128/JVI.01897-14. Epub 2014 Oct 1. J Virol. 2014. PMID: 25275125 Free PMC article.
-
Inhibition of human immunodeficiency virus Rev and human T-cell leukemia virus Rex function, but not Mason-Pfizer monkey virus constitutive transport element activity, by a mutant human nucleoporin targeted to Crm1.J Virol. 1998 Nov;72(11):8627-35. doi: 10.1128/JVI.72.11.8627-8635.1998. J Virol. 1998. PMID: 9765402 Free PMC article.
-
The export receptor Crm1 forms a dimer to promote nuclear export of HIV RNA.Elife. 2014 Dec 8;3:e04121. doi: 10.7554/eLife.04121. Elife. 2014. PMID: 25486595 Free PMC article.
-
Nucleocytoplasmic RNA transport in retroviral replication.Results Probl Cell Differ. 2001;34:197-217. doi: 10.1007/978-3-540-40025-7_12. Results Probl Cell Differ. 2001. PMID: 11288676 Review.
-
HIV-1 Rev multimerization: mechanism and insights.Curr HIV Res. 2013 Dec;11(8):623-34. doi: 10.2174/1570162x12666140307094603. Curr HIV Res. 2013. PMID: 24606219 Review.
Cited by
-
Expression, purification, and characterization of biologically active full-length Mason-Pfizer monkey virus (MPMV) Pr78Gag.Sci Rep. 2018 Aug 7;8(1):11793. doi: 10.1038/s41598-018-30142-0. Sci Rep. 2018. PMID: 30087395 Free PMC article.
-
Purification and Functional Characterization of a Biologically Active Full-Length Feline Immunodeficiency Virus (FIV) Pr50Gag.Viruses. 2019 Jul 27;11(8):689. doi: 10.3390/v11080689. Viruses. 2019. PMID: 31357656 Free PMC article.
-
HIV-1 Vif's Capacity To Manipulate the Cell Cycle Is Species Specific.J Virol. 2018 Mar 14;92(7):e02102-17. doi: 10.1128/JVI.02102-17. Print 2018 Apr 1. J Virol. 2018. PMID: 29321323 Free PMC article.
-
HIV Rev-isited.Open Biol. 2020 Dec;10(12):200320. doi: 10.1098/rsob.200320. Epub 2020 Dec 23. Open Biol. 2020. PMID: 33352061 Free PMC article. Review.
-
HIV RGB: Automated Single-Cell Analysis of HIV-1 Rev-Dependent RNA Nuclear Export and Translation Using Image Processing in KNIME.Viruses. 2022 Apr 26;14(5):903. doi: 10.3390/v14050903. Viruses. 2022. PMID: 35632645 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
