

An NK cell line (haNK) expressing high levels of granzyme and engineered to express the high affinity CD16 allele
- PMID: 27861156
- PMCID: PMC5341330
- DOI: 10.18632/oncotarget.13411
An NK cell line (haNK) expressing high levels of granzyme and engineered to express the high affinity CD16 allele
Abstract
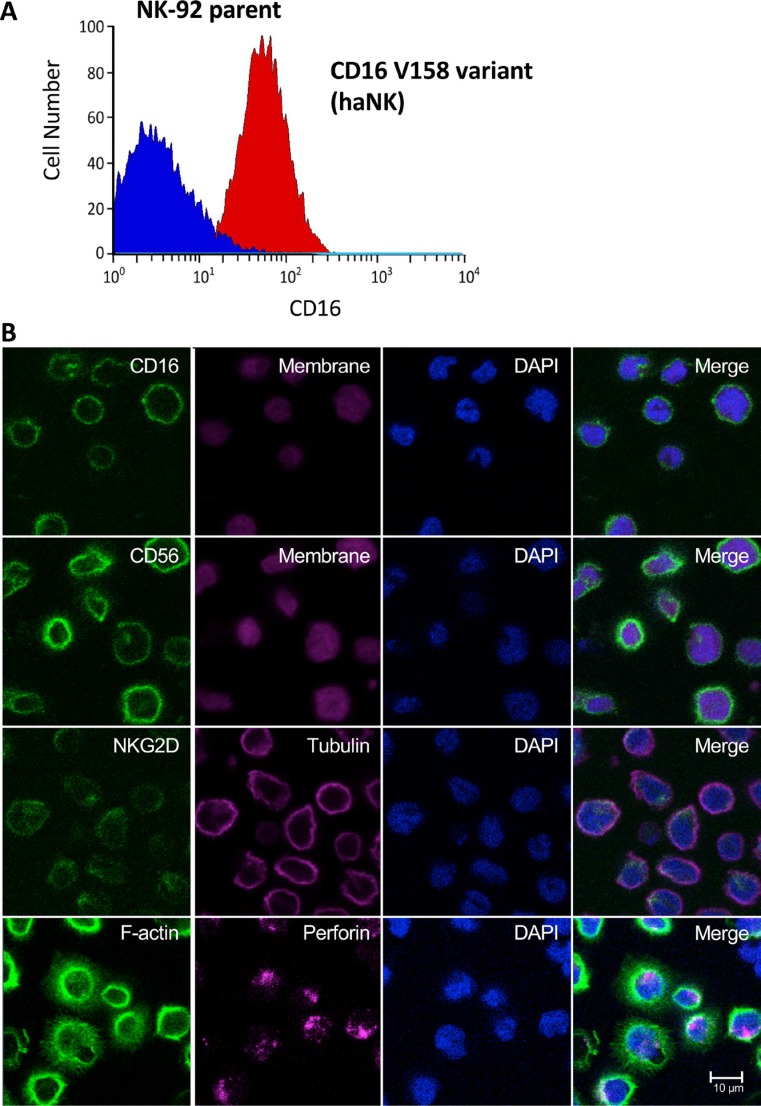
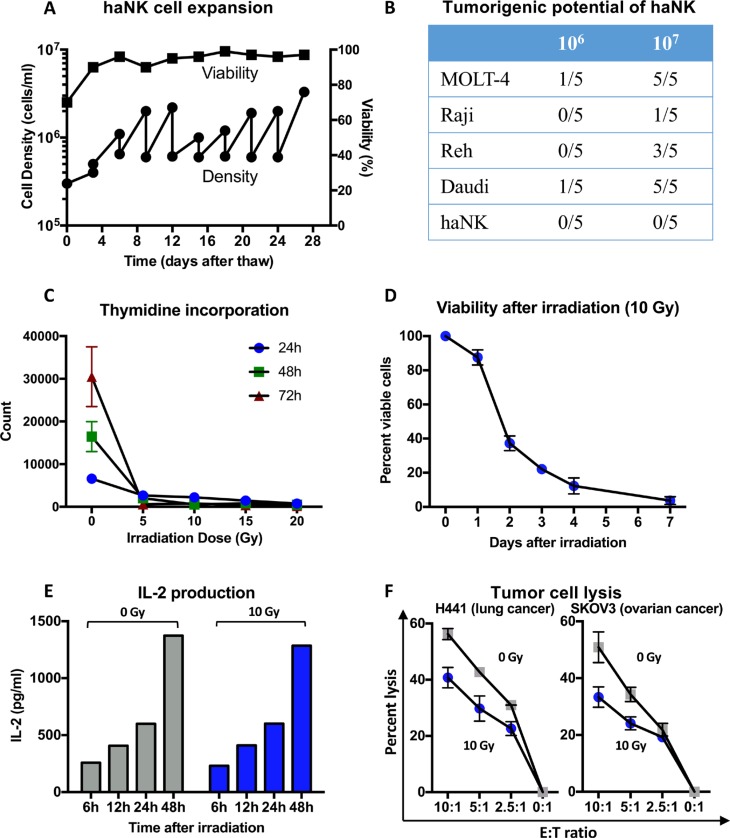
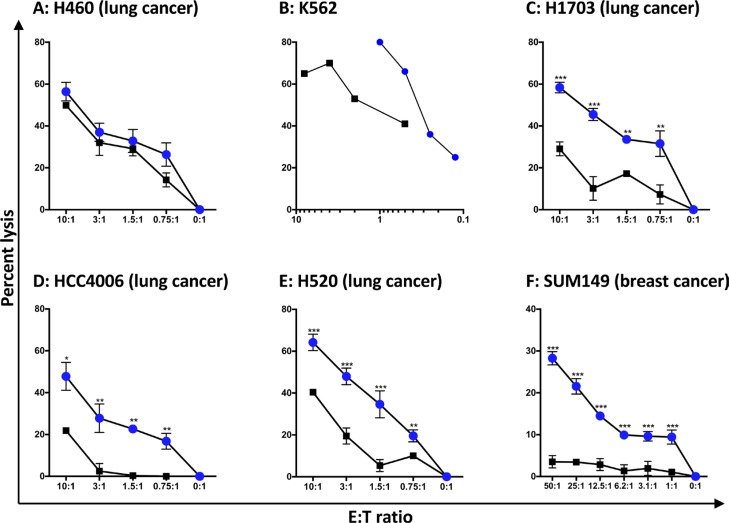
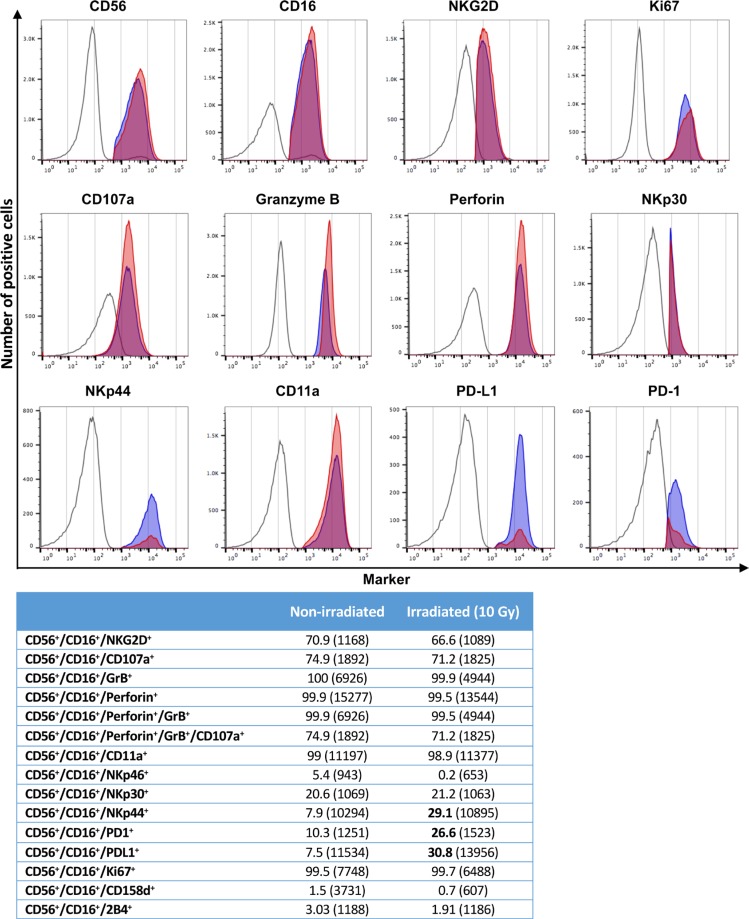
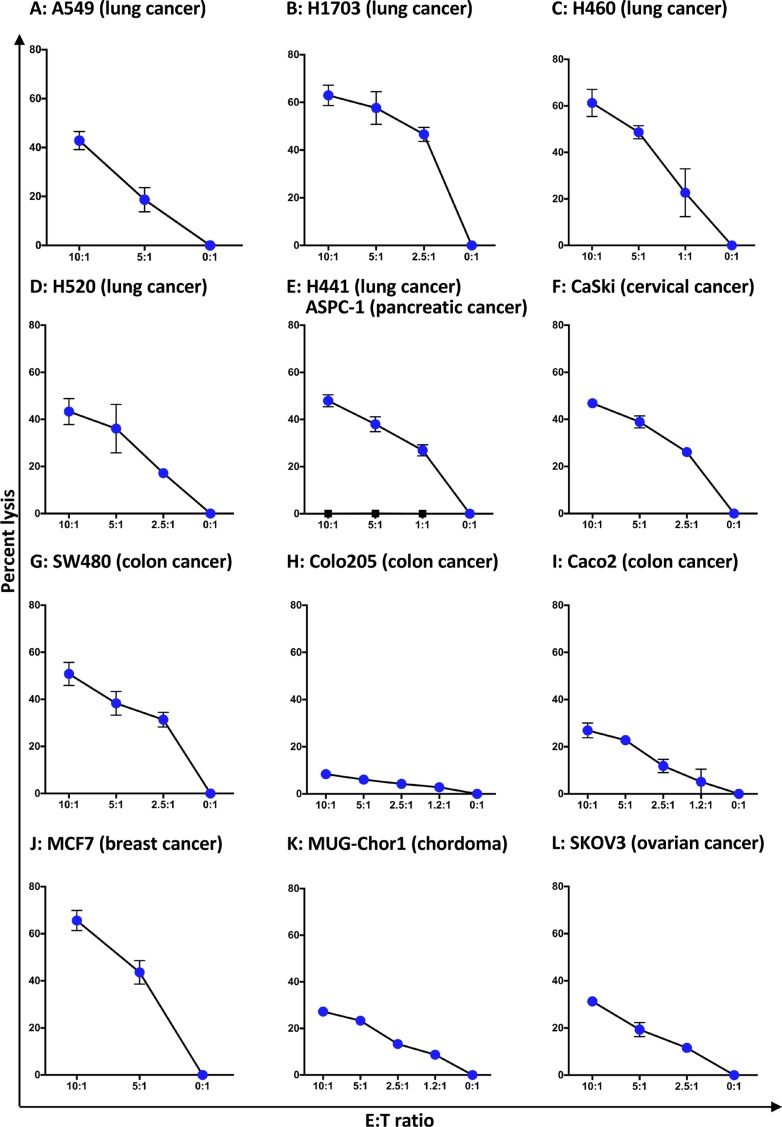
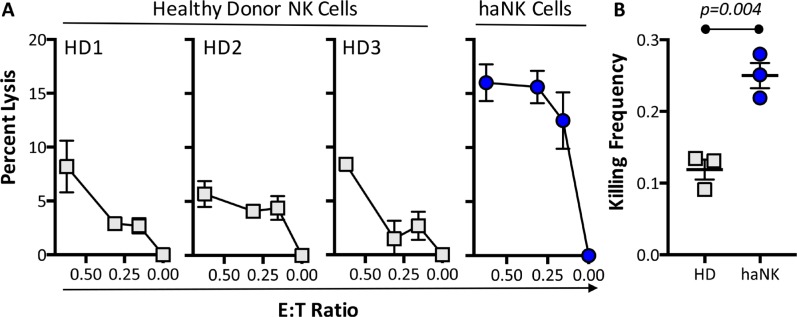
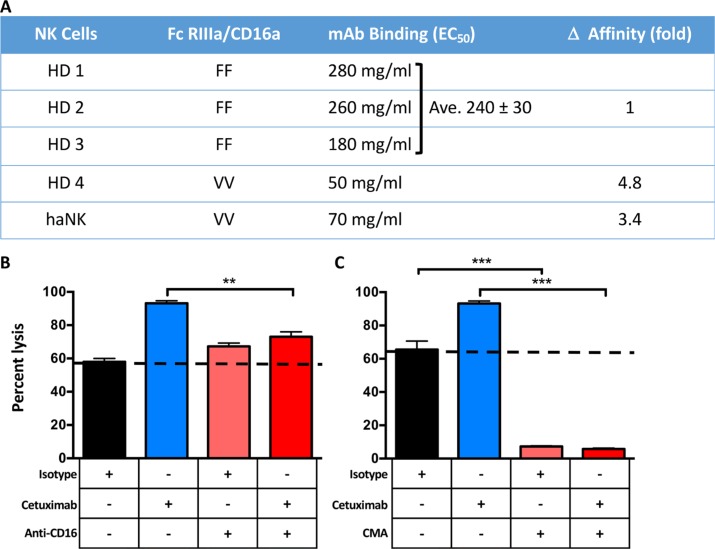
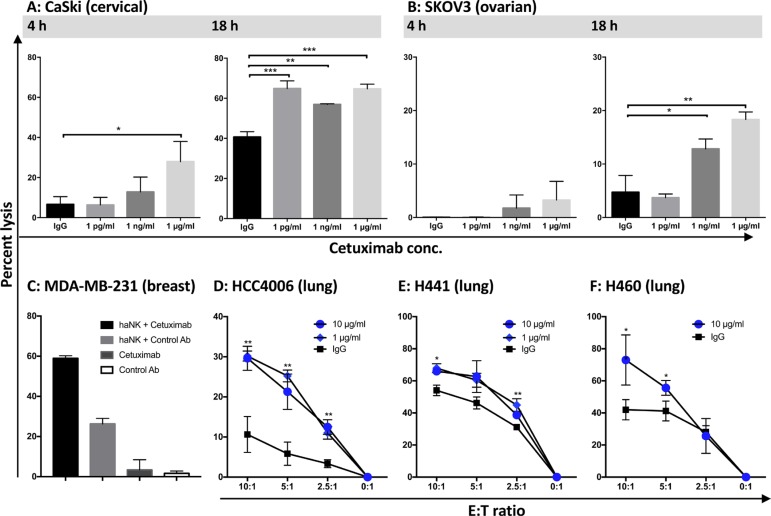
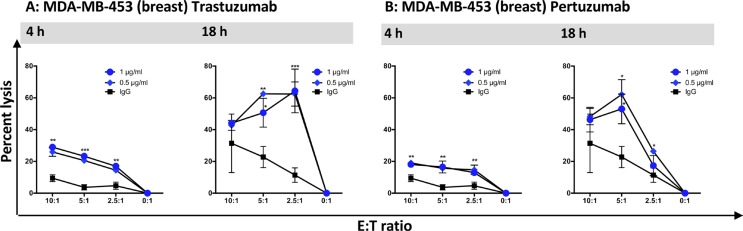
Natural killer (NK) cells are known to play a role in mediating innate immunity, in enhancing adaptive immune responses, and have been implicated in mediating anti-tumor responses via antibody-dependent cell-mediated cytotoxicity (ADCC) by reactivity of CD16 with the Fc region of human IgG1 antibodies. The NK-92 cell line, derived from a lymphoma patient, has previously been well characterized and adoptive transfer of irradiated NK-92 cells has demonstrated safety and shown preliminary evidence of clinical benefit in cancer patients. The NK-92 cell line, devoid of CD16, has now been engineered to express the high affinity (ha) CD16 V158 FcγRIIIa receptor, as well as engineered to express IL-2; IL-2 has been shown to replenish the granular stock of NK cells, leading to enhanced perforin- and granzyme-mediated lysis of tumor cells. The studies reported here show high levels of granzyme in haNK cells, and demonstrate the effects of irradiation of haNK cells on multiple phenotypic markers, viability, IL-2 production, and lysis of a spectrum of human tumor cells. Studies also compare endogenous irradiated haNK lysis of tumor cells with that of irradiated haNK-mediated ADCC using cetuximab, trastuzumab and pertuzumab monoclonal antibodies. These studies thus provide the rationale for the potential use of irradiated haNK cells in adoptive transfer studies for a range of human tumor types. Moreover, since only approximately 10% of humans are homozygous for the high affinity V CD16 allele, these studies also provide the rationale for the use of irradiated haNK cells in combination with IgG1 anti-tumor monoclonal antibodies.
Keywords: ADCC; NK lysis; cetuximab; high affinity CD16; immunotherapy.
Conflict of interest statement
The authors have no potential conflicts of interest to disclose. The Laboratory of Tumor Immunology and Biology, National Cancer Institute, has a Cooperative Research and Development Agreement (CRADA) with NantBioScience.
No potential conflicts of interest to disclose.
Hans Klingemann and Laurent Boissel are employees of NantKwest. Patrick Soon-Shiong is a founder and an executive at NantKwest and NantCell, Shahrooz Rabizadeh is a contributor to NantCell.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
