

Ever-Changing Landscapes: Transcriptional Enhancers in Development and Evolution
- PMID: 27863239
- PMCID: PMC5123704
- DOI: 10.1016/j.cell.2016.09.018
Ever-Changing Landscapes: Transcriptional Enhancers in Development and Evolution
Abstract
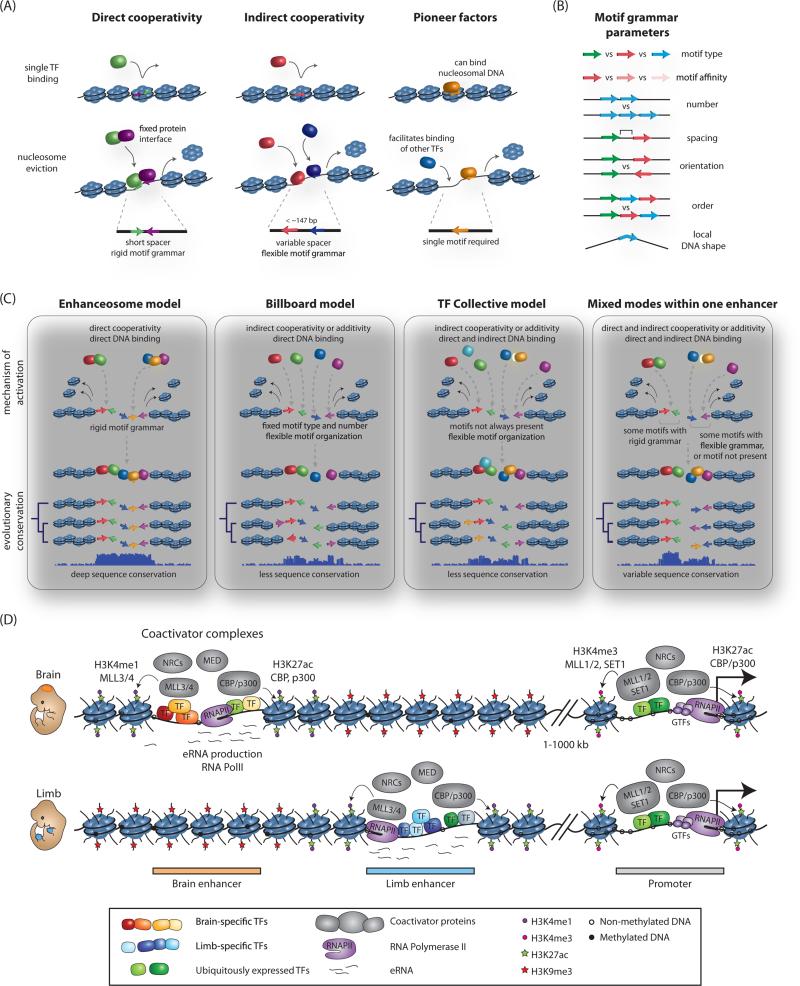
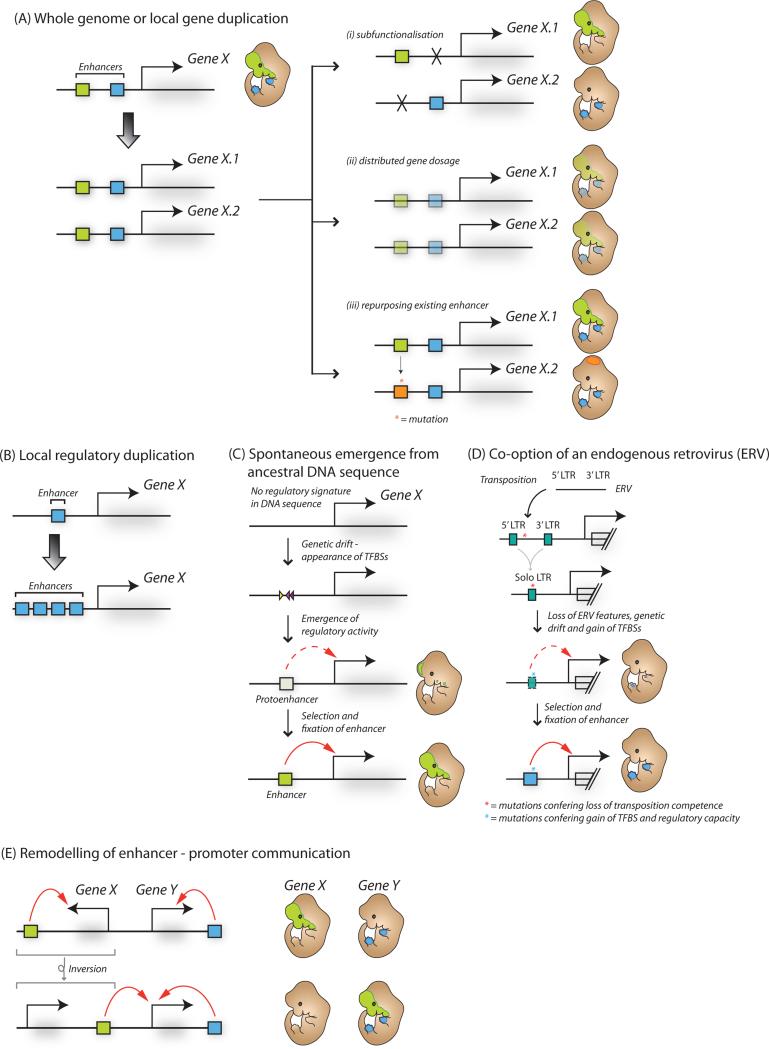
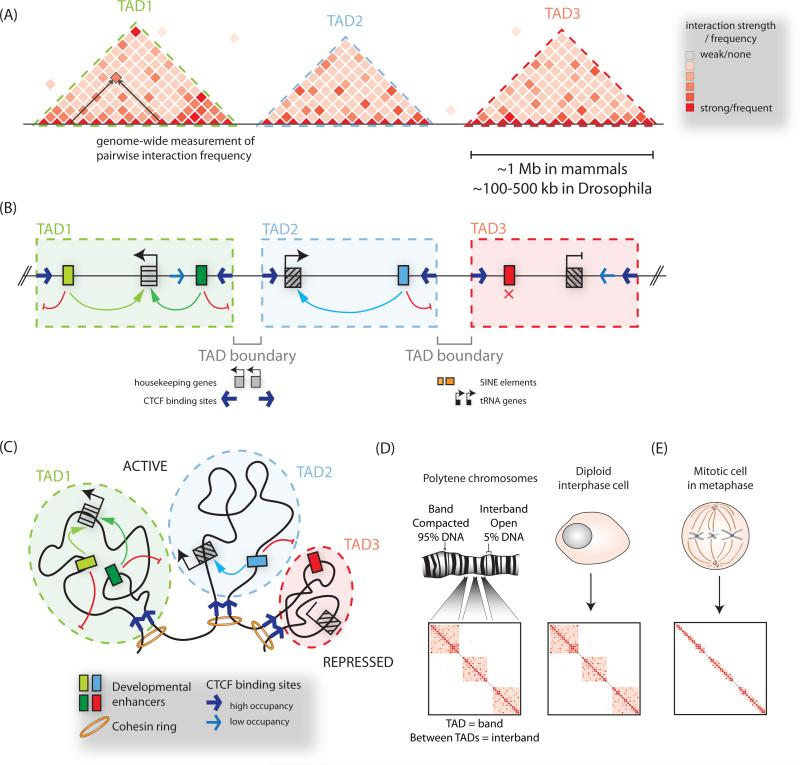
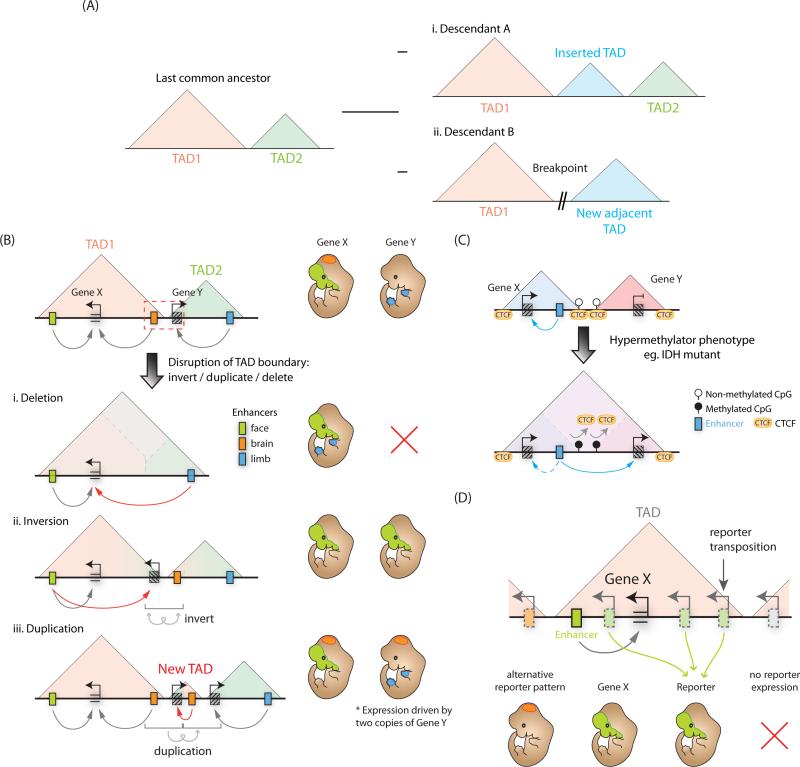
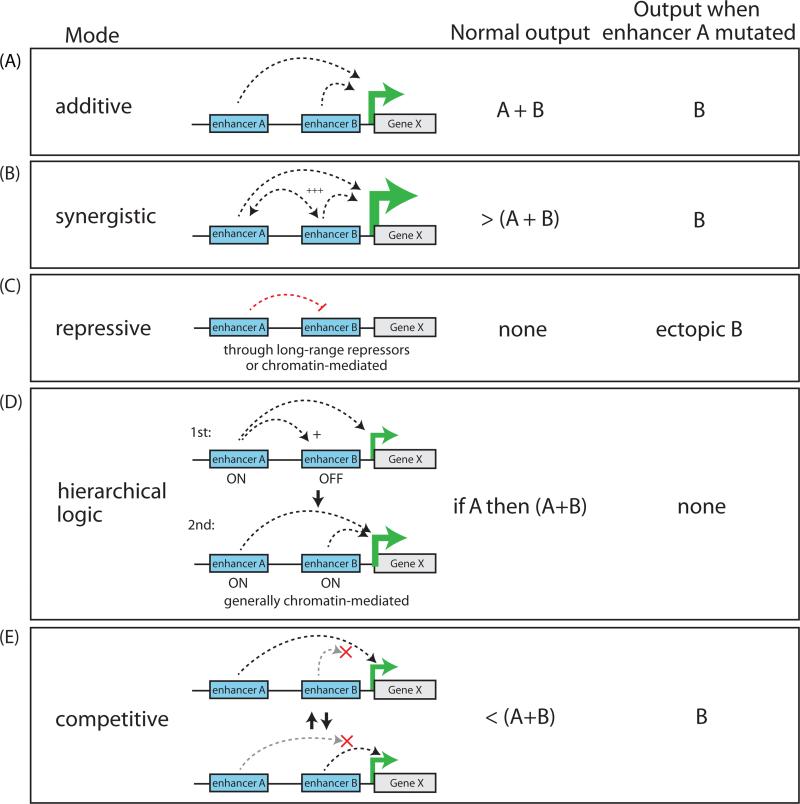
A class of cis-regulatory elements, called enhancers, play a central role in orchestrating spatiotemporally precise gene-expression programs during development. Consequently, divergence in enhancer sequence and activity is thought to be an important mediator of inter- and intra-species phenotypic variation. Here, we give an overview of emerging principles of enhancer function, current models of enhancer architecture, genomic substrates from which enhancers emerge during evolution, and the influence of three-dimensional genome organization on long-range gene regulation. We discuss intricate relationships between distinct elements within complex regulatory landscapes and consider their potential impact on specificity and robustness of transcriptional regulation.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Allan CM, Walker D, Taylor JM. Evolutionary duplication of a hepatic control region in the human apolipoprotein E gene locus. Identification of a second region that confers high level and liver-specific expression of the human apolipoprotein E gene in transgenic mice. J. Biol. Chem. 1995;270:26278–26281. - PubMed
-
- Arnosti DN, Kulkarni MM. Transcriptional enhancers: Intelligent enhanceosomes or flexible billboards? J. Cell. Biochem. 2005;94:890–898. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
