

Lipid zonation and phospholipid remodeling in nonalcoholic fatty liver disease
- PMID: 27863448
- PMCID: PMC5396354
- DOI: 10.1002/hep.28953
Lipid zonation and phospholipid remodeling in nonalcoholic fatty liver disease
Abstract
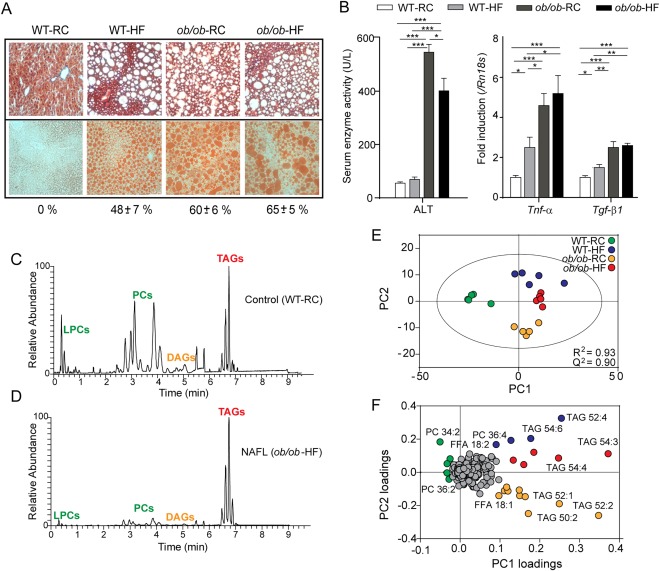
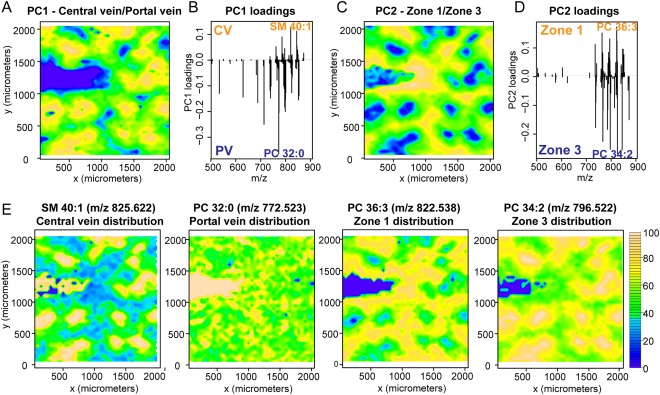
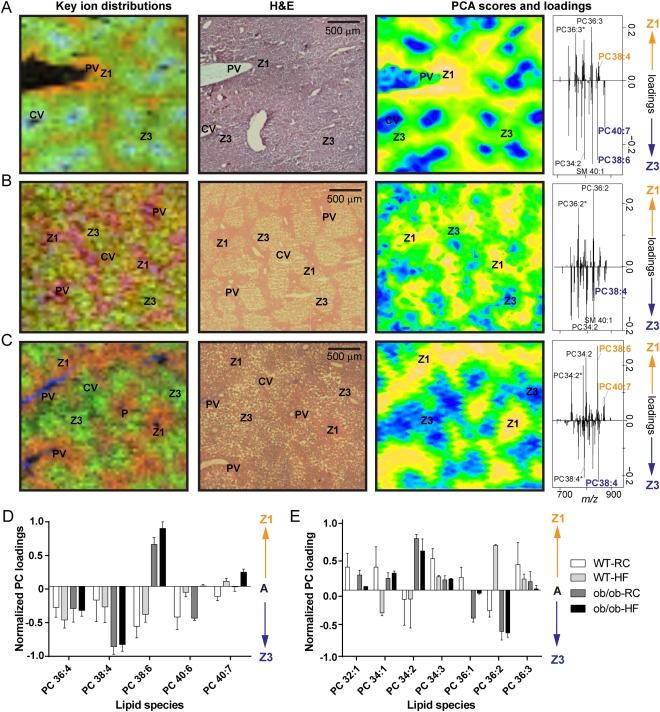
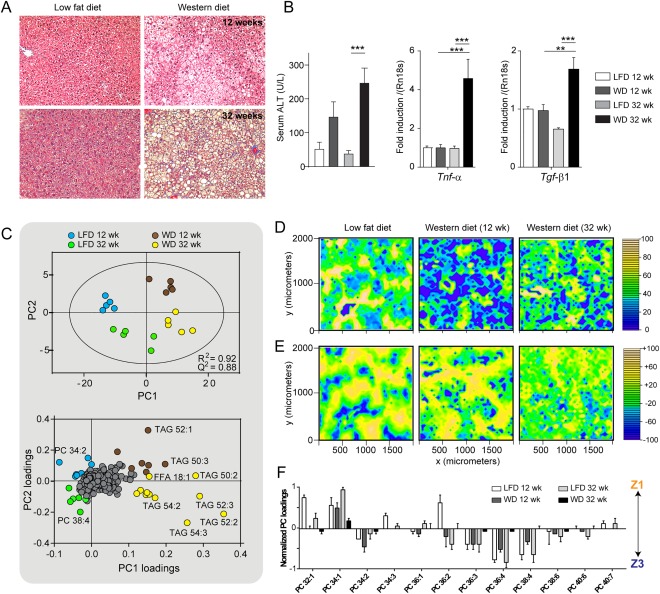
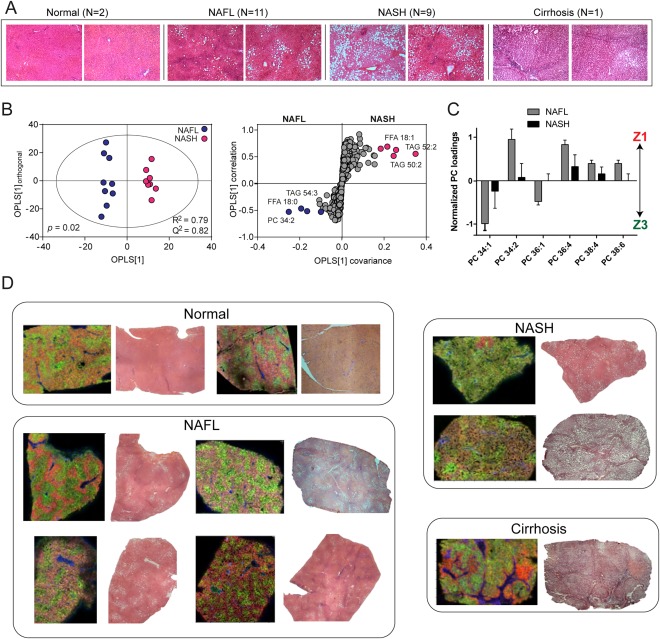
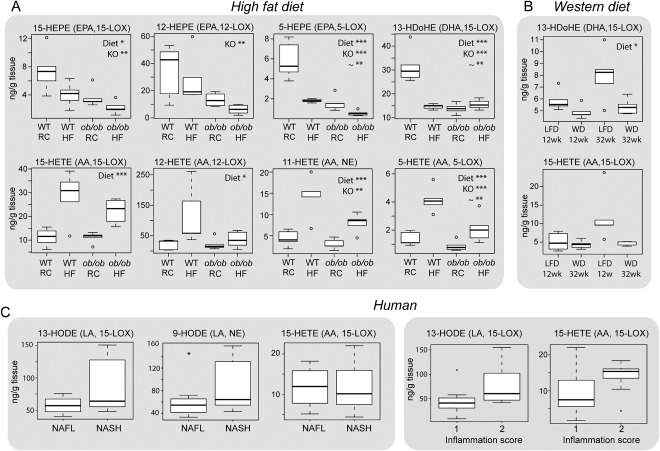
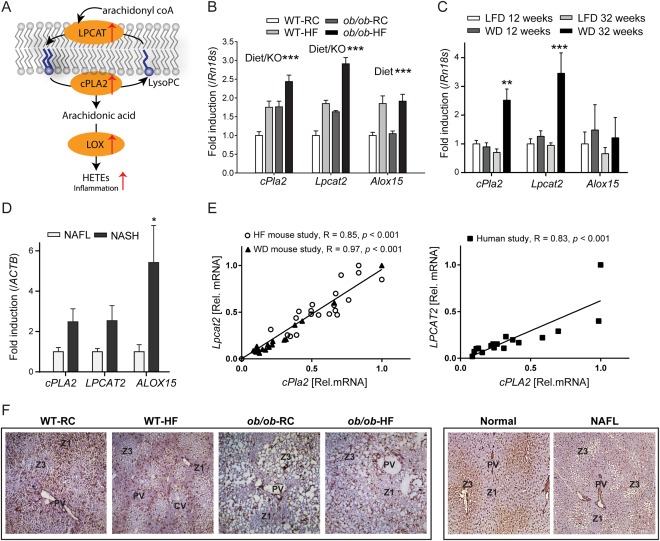
Nonalcoholic fatty liver disease (NAFLD) can progress from simple steatosis (i.e., nonalcoholic fatty liver [NAFL]) to nonalcoholic steatohepatitis (NASH), cirrhosis, and cancer. Currently, the driver for this progression is not fully understood; in particular, it is not known how NAFLD and its early progression affects the distribution of lipids in the liver, producing lipotoxicity and inflammation. In this study, we used dietary and genetic mouse models of NAFL and NASH and translated the results to humans by correlating the spatial distribution of lipids in liver tissue with disease progression using advanced mass spectrometry imaging technology. We identified several lipids with distinct zonal distributions in control and NAFL samples and observed partial to complete loss of lipid zonation in NASH. In addition, we found increased hepatic expression of genes associated with remodeling the phospholipid membrane, release of arachidonic acid (AA) from the membrane, and production of eicosanoid species that promote inflammation and cell injury. The results of our immunohistochemistry analyses suggest that the zonal location of remodeling enzyme LPCAT2 plays a role in the change in spatial distribution for AA-containing lipids. This results in a cycle of AA-enrichment in pericentral hepatocytes, membrane release of AA, and generation of proinflammatory eicosanoids and may account for increased oxidative damage in pericentral regions in NASH.
Conclusion: NAFLD is associated not only with lipid enrichment, but also with zonal changes of specific lipids and their associated metabolic pathways. This may play a role in the heterogeneous development of NAFLD. (Hepatology 2017;65:1165-1180).
© 2016 by The Authors. Hepatology published by Wiley Periodicals, Inc., on behalf of the American Association for the Study of Liver Diseases.
Figures
References
-
- Yeh MM, Brunt EM. Pathological features of fatty liver disease. Gastroenterology 2014;147:754‐764. - PubMed
-
- Wree A, Broderick L, Canbay A, Hoffman HM, Feldstein AE. From NAFLD to NASH to cirrhosis‐new insights into disease mechanisms. Nat Rev Gastroenterol Hepatol 2013;10:627‐636. - PubMed
-
- Vacca M, Allison M, Griffin JL, Vidal‐Puig A. Fatty acid and glucose sensors in hepatic lipid metabolism: implications in NAFLD. Semin Liver Dis 2015;35:250‐261. - PubMed
-
- Tilg H, Moschen AR. Evolution of inflammation in nonalcoholic fatty liver disease: the multiple parallel hits hypothesis. Hepatology 2010;52:1836‐1846. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- MC_G0802535/MRC_/Medical Research Council/United Kingdom
- MC_UU_12012/2/MRC_/Medical Research Council/United Kingdom
- BB/H002731/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G0600717/MRC_/Medical Research Council/United Kingdom
- MC_UP_A090_1006/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
