

Root nodule symbiosis in Lotus japonicus drives the establishment of distinctive rhizosphere, root, and nodule bacterial communities
- PMID: 27864511
- PMCID: PMC5150415
- DOI: 10.1073/pnas.1616564113
Root nodule symbiosis in Lotus japonicus drives the establishment of distinctive rhizosphere, root, and nodule bacterial communities
Abstract
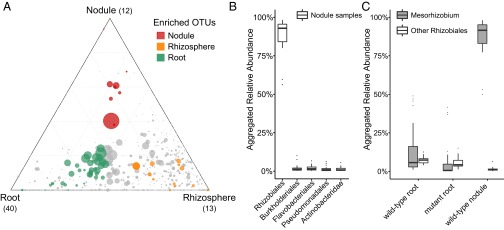
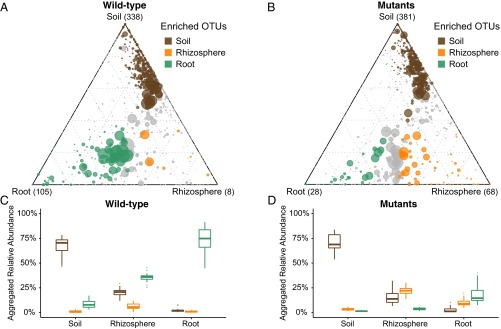
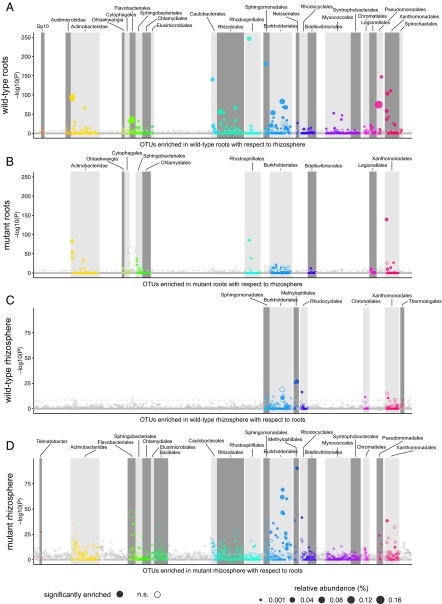
Lotus japonicus has been used for decades as a model legume to study the establishment of binary symbiotic relationships with nitrogen-fixing rhizobia that trigger root nodule organogenesis for bacterial accommodation. Using community profiling of 16S rRNA gene amplicons, we reveal that in Lotus, distinctive nodule- and root-inhabiting communities are established by parallel, rather than consecutive, selection of bacteria from the rhizosphere and root compartments. Comparative analyses of wild-type (WT) and symbiotic mutants in Nod factor receptor5 (nfr5), Nodule inception (nin) and Lotus histidine kinase1 (lhk1) genes identified a previously unsuspected role of the nodulation pathway in the establishment of different bacterial assemblages in the root and rhizosphere. We found that the loss of nitrogen-fixing symbiosis dramatically alters community structure in the latter two compartments, affecting at least 14 bacterial orders. The differential plant growth phenotypes seen between WT and the symbiotic mutants in nonsupplemented soil were retained under nitrogen-supplemented conditions that blocked the formation of functional nodules in WT, whereas the symbiosis-impaired mutants maintain an altered community structure in the nitrogen-supplemented soil. This finding provides strong evidence that the root-associated community shift in the symbiotic mutants is a direct consequence of the disabled symbiosis pathway rather than an indirect effect resulting from abolished symbiotic nitrogen fixation. Our findings imply a role of the legume host in selecting a broad taxonomic range of root-associated bacteria that, in addition to rhizobia, likely contribute to plant growth and ecological performance.
Keywords: 16S; Lotus japonicus; microbiota; nitrogen fixation; symbiosis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
Lotus japonicus Symbiosis Genes Impact Microbial Interactions between Symbionts and Multikingdom Commensal Communities.mBio. 2019 Oct 8;10(5):e01833-19. doi: 10.1128/mBio.01833-19. mBio. 2019. PMID: 31594815 Free PMC article.
-
Spontaneous symbiotic reprogramming of plant roots triggered by receptor-like kinases.Elife. 2014 Nov 25;3:e03891. doi: 10.7554/eLife.03891. Elife. 2014. PMID: 25422918 Free PMC article.
-
Loss-of-function of ASPARTIC PEPTIDASE NODULE-INDUCED 1 (APN1) in Lotus japonicus restricts efficient nitrogen-fixing symbiosis with specific Mesorhizobium loti strains.Plant J. 2018 Jan;93(1):5-16. doi: 10.1111/tpj.13759. Epub 2017 Dec 2. Plant J. 2018. PMID: 29086445
-
Leguminous plants: inventors of root nodules to accommodate symbiotic bacteria.Int Rev Cell Mol Biol. 2015;316:111-58. doi: 10.1016/bs.ircmb.2015.01.004. Epub 2015 Feb 20. Int Rev Cell Mol Biol. 2015. PMID: 25805123 Review.
-
Transcription factors network in root endosymbiosis establishment and development.World J Microbiol Biotechnol. 2018 Feb 15;34(3):37. doi: 10.1007/s11274-018-2418-7. World J Microbiol Biotechnol. 2018. PMID: 29450655 Review.
Cited by
-
Nodulation and nitrogen fixation in Medicago truncatula strongly alters the abundance of its root microbiota and subtly affects its structure.Environ Microbiol. 2022 Nov;24(11):5524-5533. doi: 10.1111/1462-2920.16164. Epub 2022 Aug 31. Environ Microbiol. 2022. PMID: 36054464 Free PMC article.
-
Host genetic determinants drive compartment-specific assembly of tea plant microbiomes.Plant Biotechnol J. 2022 Nov;20(11):2174-2186. doi: 10.1111/pbi.13897. Epub 2022 Aug 8. Plant Biotechnol J. 2022. PMID: 35876474 Free PMC article.
-
Trypoxylus dichotomus Gut Bacteria Provides an Effective System for Bamboo Lignocellulose Degradation.Microbiol Spectr. 2022 Oct 26;10(5):e0214722. doi: 10.1128/spectrum.02147-22. Epub 2022 Aug 22. Microbiol Spectr. 2022. PMID: 35993784 Free PMC article.
-
Nodulation number tempers the relative importance of stochastic processes in the assembly of soybean root-associated communities.ISME Commun. 2023 Aug 28;3(1):89. doi: 10.1038/s43705-023-00296-8. ISME Commun. 2023. PMID: 37640896 Free PMC article.
-
Variation in rhizosphere microbial communities and its association with the symbiotic efficiency of rhizobia in soybean.ISME J. 2020 Aug;14(8):1915-1928. doi: 10.1038/s41396-020-0648-9. Epub 2020 Apr 27. ISME J. 2020. PMID: 32336748 Free PMC article.
References
-
- Maathuis FJ. Physiological functions of mineral macronutrients. Curr Opin Plant Biol. 2009;12(3):250–258. - PubMed
-
- Bulgarelli D, Schlaeppi K, Spaepen S, Ver Loren van Themaat E, Schulze-Lefert P. Structure and functions of the bacterial microbiota of plants. Annu Rev Plant Biol. 2013;64:807–838. - PubMed
-
- Parniske M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat Rev Microbiol. 2008;6(10):763–775. - PubMed
-
- Vorholt JA. Microbial life in the phyllosphere. Nat Rev Microbiol. 2012;10(12):828–840. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
