

Review
doi: 10.1016/j.cell.2016.10.044.
Epub 2016 Nov 17.
CRISPR-Based Technologies for the Manipulation of Eukaryotic Genomes
Affiliations
- PMID: 27866654
- PMCID: PMC5235943
- DOI: 10.1016/j.cell.2016.10.044
Item in Clipboard
Review
CRISPR-Based Technologies for the Manipulation of Eukaryotic Genomes
Cell.
.
Erratum in
-
CRISPR-Based Technologies for the Manipulation of Eukaryotic Genomes.Cell. 2017 Apr 20;169(3):559. doi: 10.1016/j.cell.2017.04.005. Cell. 2017. PMID: 28431253 No abstract available.
Abstract
The CRISPR-Cas9 RNA-guided DNA endonuclease has contributed to an explosion of advances in the life sciences that have grown from the ability to edit genomes within living cells. In this Review, we summarize CRISPR-based technologies that enable mammalian genome editing and their various applications. We describe recent developments that extend the generality, DNA specificity, product selectivity, and fundamental capabilities of natural CRISPR systems, and we highlight some of the remarkable advancements in basic research, biotechnology, and therapeutics science that these developments have facilitated.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
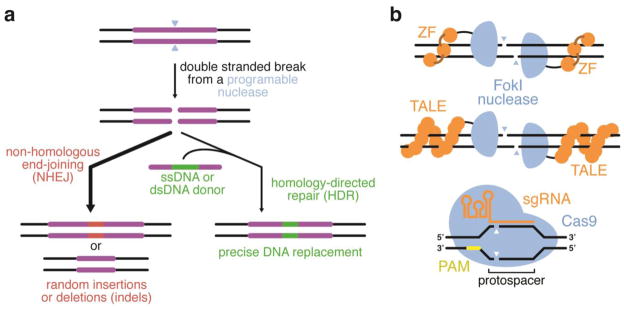
Genome editing using double-stranded breaks (DSBs). (a) A programmable nuclease incorporates a sequence-specific DSB in genomic DNA. In the absence of a repair template, the cell will process the DSB mostly by NHEJ, resulting in indels at the site of editing. In the presence of a separate DNA template containing sequences homologous to the regions flanking the DSB, HDR can result in incorporation of the repair template into the genomic DNA. (b) ZFNs, TALENs, and CRISPR-based nucleases have also been used to introduce programmable, sequence-specific DSBs. The ability of Cas9 to be reprogrammed to bind a new 23-bp sequence (the protospacer and PAM) by designing a new sgRNA, rather than by engineering a new DNA-binding protein (orange), has transformed the genome editing field.
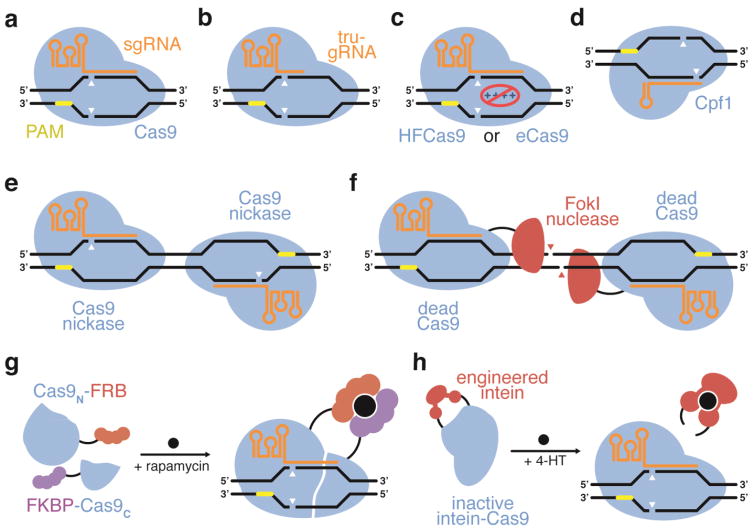
Strategies for improving the DNA specificity of CRISPR-based agents. (a) Wild-type Cas9 variants have been shown to possess significant off-target activity. (b) DNA specificity can be improved using truncated sgRNAs with wtCas9 (Fu et al., 2014), (c) engineered HFCas9 or eCas9 variants that reduce nonspecific electrostatic interactions between the protein and DNA (Slaymaker et al., 2016; Kleinstiver et al., 2016a), or (d) the Cpf1 CRISPR enzyme (Kim et al., 2016; Kleinstiver et al., 2016b). Alternatively, (e) two Cas9 nickase enzymes (Ran et al., 2013a; Mali et al., 2013a), or (f) dCas9-FokI fusions can be used to require two RNA-programmed binding events to induce a DSB (Guilinger et al., 2014b), increasing specificity. DNA specificity can also be increased by limiting the cellular residence time of wtCas9 using (g) a small molecule-activated split Cas9 (Zetsche et al., 2015b), or (h) a small molecule-activated intein-disrupted Cas9 (Davis et al., 2015).
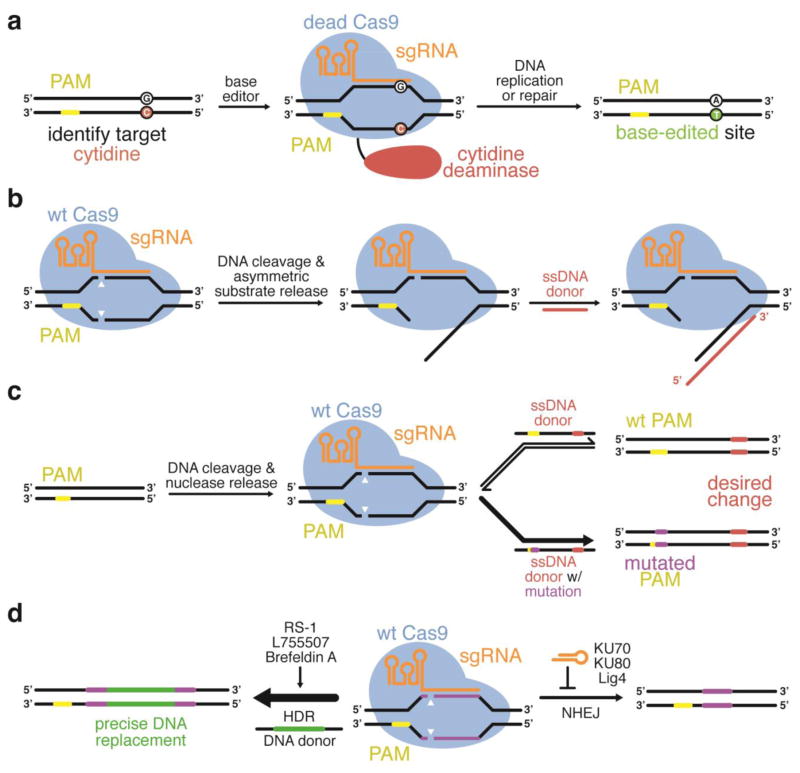
Approaches that improve the product selectivity of genome editing agents. Wild-type Cas9 will induce undesired indels when the desired product is a precise DNA modification. (a) Base editing is capable of editing G:C base pairs to A:T base pairs with high conversions and very low indel rates (Nishida et al., 2016; Komor et al., 2016). (b) The ssDNA donor used during HDR can be designed such that it anneals with the DNA strand that is initially released by Cas9 following DNA cleavage to enhance HDR efficiency (Richardson et al., 2016). (c) In some cases, HDR strategies can also be designed to install a silent mutation into the PAM in order to prevent re-cutting by Cas9 following HDR (Paquet et al., 2016). (d) Small molecule inhibitors of NHEJ (Srivastava et al., 2012; Robert et al., 2015; Vartak and Raghavan, 2015; Chu et al., 2015; Maruyama et al., 2015), enhancers of HDR (Yu et al., 2015; Pinder et al., 2015; Song et al., 2016), or cell-cycle synchronizers (Lin et al., 2014b) can be used to increase the ratio of HDR:NHEJ genome editing products.
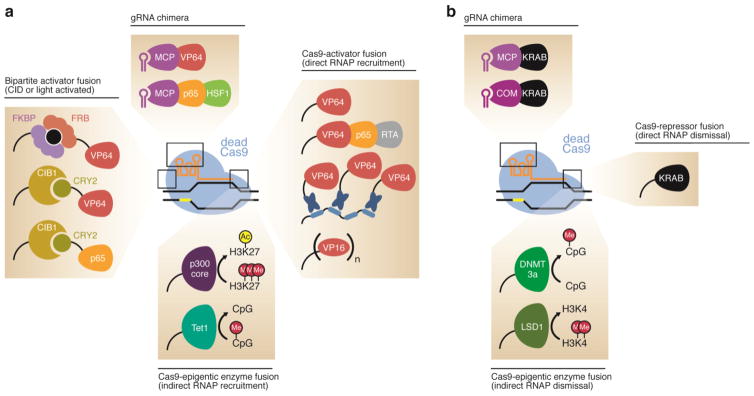
CRISPR-based epigenome editing. (a) RNA-programmed gene activators can be assembled through the direct fusion of dCas9 with the transcriptional activators VP64 (Mali et al., 2013a; Maeder et al., 2013; Perez-Pinera et al., 2013; Tanenbaum et al., 2014) and VPR (Chavez et al., 2015), the histone acetyltransferase enzyme p300 (Hilton et al., 2015), or the DNA demethylase Tet1 (Xu et al., 2016; Morita et al., 2016; Liu et al., 2016). Alternately, the transcriptional activators VP64 and p65-HSF1 can be attached to the sgRNA (Zalatan et al., 2015; Konermann et al., 2015). Light-activated and small molecule-activated variants can also be used (Nihongaki et al., 2015b; Zetsche et al., 2015b; Polstein and Gersbach, 2015). (b) RNA-programmed gene repressors can be assembled by attaching the transcriptional repressor domain KRAB to dCas9 by either a direct fusion between the two proteins (Gilbert et al., 2013; Lawhorn et al., 2014; Thakore et al., 2015) or via the sgRNA. Alternatively, dCas9 can be fused to the DNA methyltransferase enzyme DNMT3a (McDonald et al., 2016; Liu et al., 2016; Vojta et al., 2016) or the histone demethylase LSD1 (Kearns et al., 2015) to result in a transcriptional repressor.
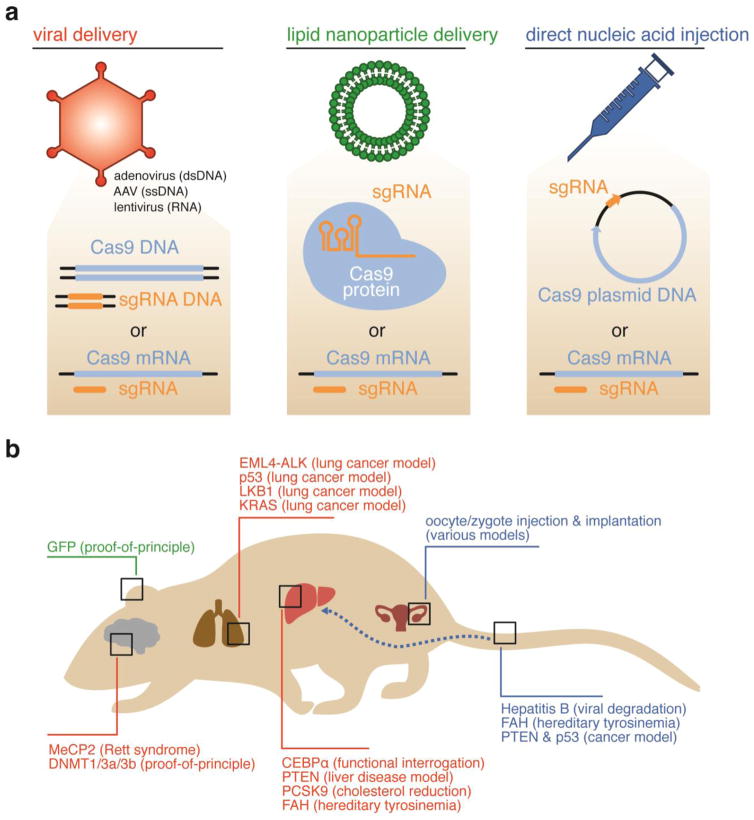
Strategies for in vivo delivery of CRISPR-based genome editing agents. (a) Viral (orange)-, lipid nanoparticle (green)-, and direct nucleic acid injection (blue)-mediated delivery of CRISPR-based genome editing agents have all been successfully used to achieve in vivo genome editing. (b) These methods have been used to deliver genome editing agents to a variety of mammalian organs shown. The genes that were modified within each organ are shown in a color corresponding to the delivery method used, matching the colors in (a).
References
-
- Amsellem S, Pflumio F, Bardinet D, Izac B, Charneau P, Romeo PH, Dubart-Kupperschmitt A, Fichelson S. Ex vivo expansion of human hematopoietic stem cells by direct delivery of the HOXB4 homeoprotein. Nat Med. 2003;9:1423–1427. - PubMed
-
- Barrangou R, Fremaux C, Deveau H, Richards M, Boyaval P, Moineau S, Romero DA, Horvath P. CRISPR Provides Acquired Resistance Against Viruses in Prokaryotes. Science. 2007;315:1709. - PubMed
-
- Bartus RT, Tracy MA, Emerich DF, Zale SE. Sustained Delivery of Proteins for Novel Therapeutic Products. Science. 1998;281:1161–1162. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
