

The HECT domain ubiquitin ligase HUWE1 targets unassembled soluble proteins for degradation
- PMID: 27867533
- PMCID: PMC5102030
- DOI: 10.1038/celldisc.2016.40
The HECT domain ubiquitin ligase HUWE1 targets unassembled soluble proteins for degradation
Abstract
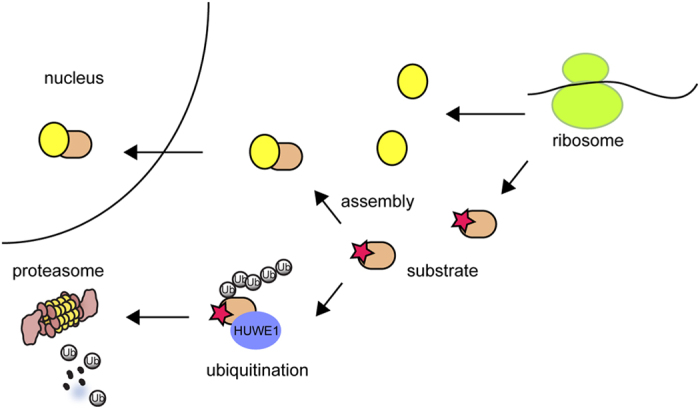
In eukaryotes, many proteins function in multi-subunit complexes that require proper assembly. To maintain complex stoichiometry, cells use the endoplasmic reticulum-associated degradation system to degrade unassembled membrane subunits, but how unassembled soluble proteins are eliminated is undefined. Here we show that degradation of unassembled soluble proteins (referred to as unassembled soluble protein degradation, USPD) requires the ubiquitin selective chaperone p97, its co-factor nuclear protein localization protein 4 (Npl4), and the proteasome. At the ubiquitin ligase level, the previously identified protein quality control ligase UBR1 (ubiquitin protein ligase E3 component n-recognin 1) and the related enzymes only process a subset of unassembled soluble proteins. We identify the homologous to the E6-AP carboxyl terminus (homologous to the E6-AP carboxyl terminus) domain-containing protein HUWE1 as a ubiquitin ligase for substrates bearing unshielded, hydrophobic segments. We used a stable isotope labeling with amino acids-based proteomic approach to identify endogenous HUWE1 substrates. Interestingly, many HUWE1 substrates form multi-protein complexes that function in the nucleus although HUWE1 itself is cytoplasmically localized. Inhibition of nuclear entry enhances HUWE1-mediated ubiquitination and degradation, suggesting that USPD occurs primarily in the cytoplasm. Altogether, these findings establish a new branch of the cytosolic protein quality control network, which removes surplus polypeptides to control protein homeostasis and nuclear complex assembly.
Keywords: HUWE1; p97/Cdc48; protein quality control (PQC); ubiquitin; unassembled soluble protein degradation (USPD).
Figures
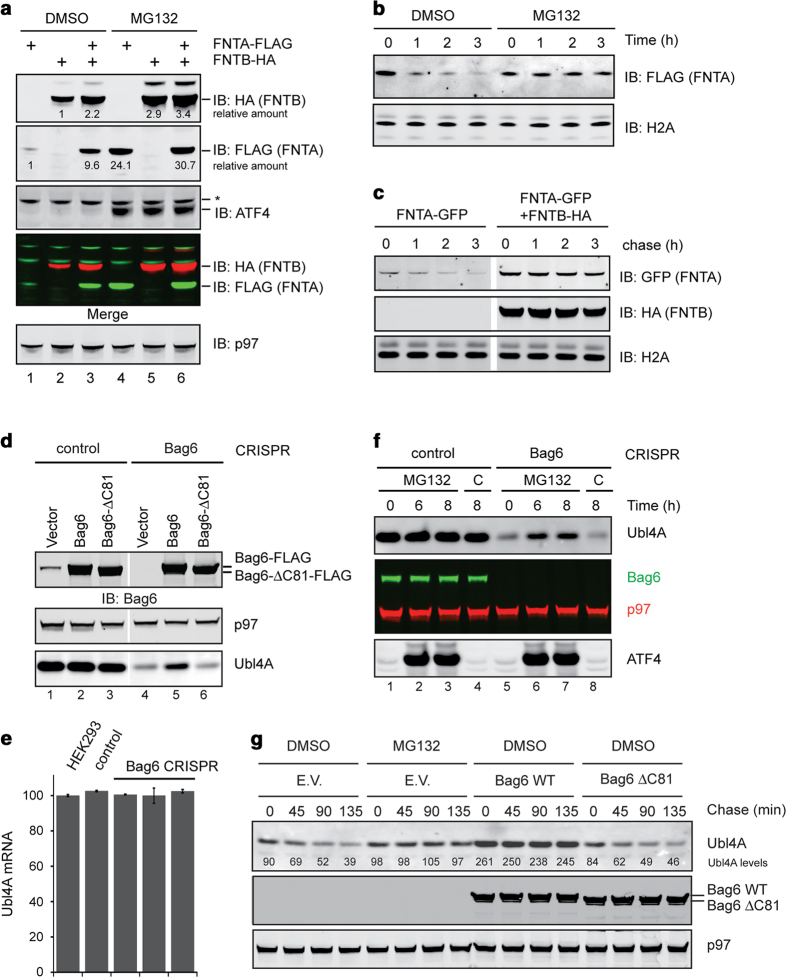


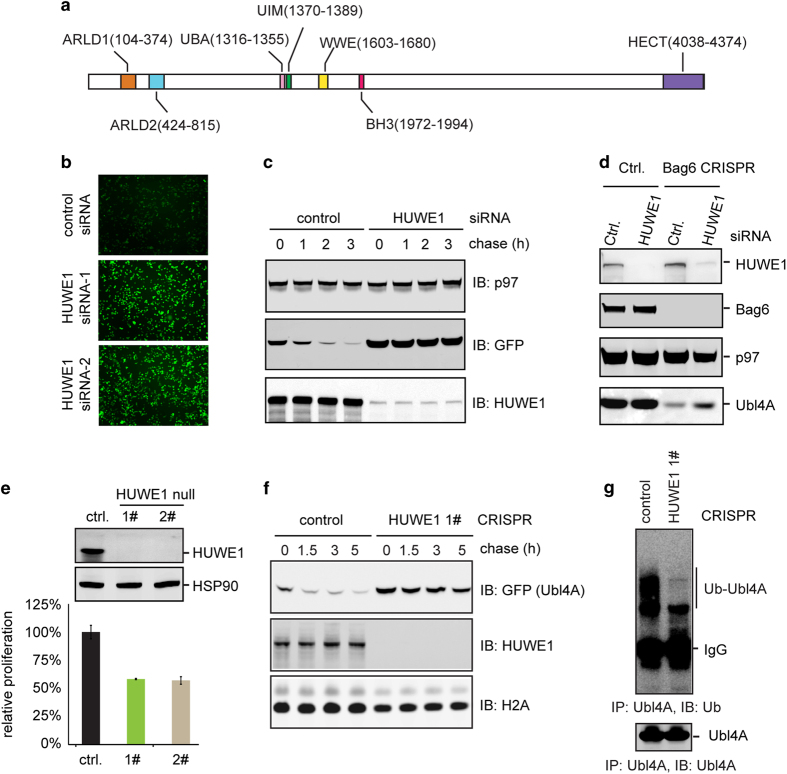
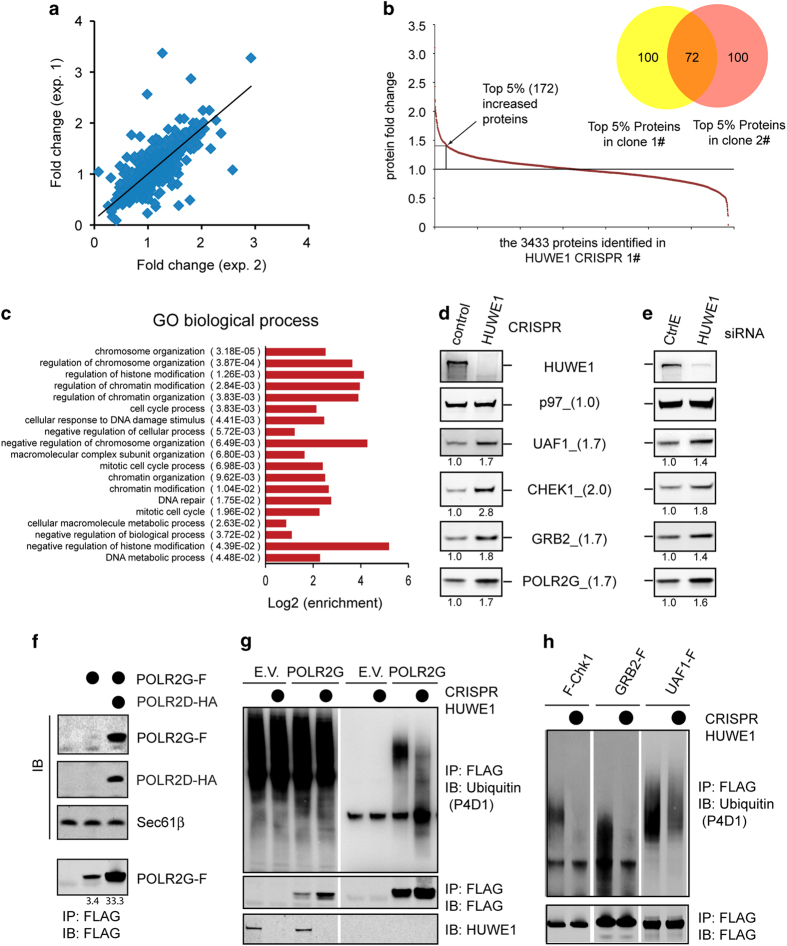
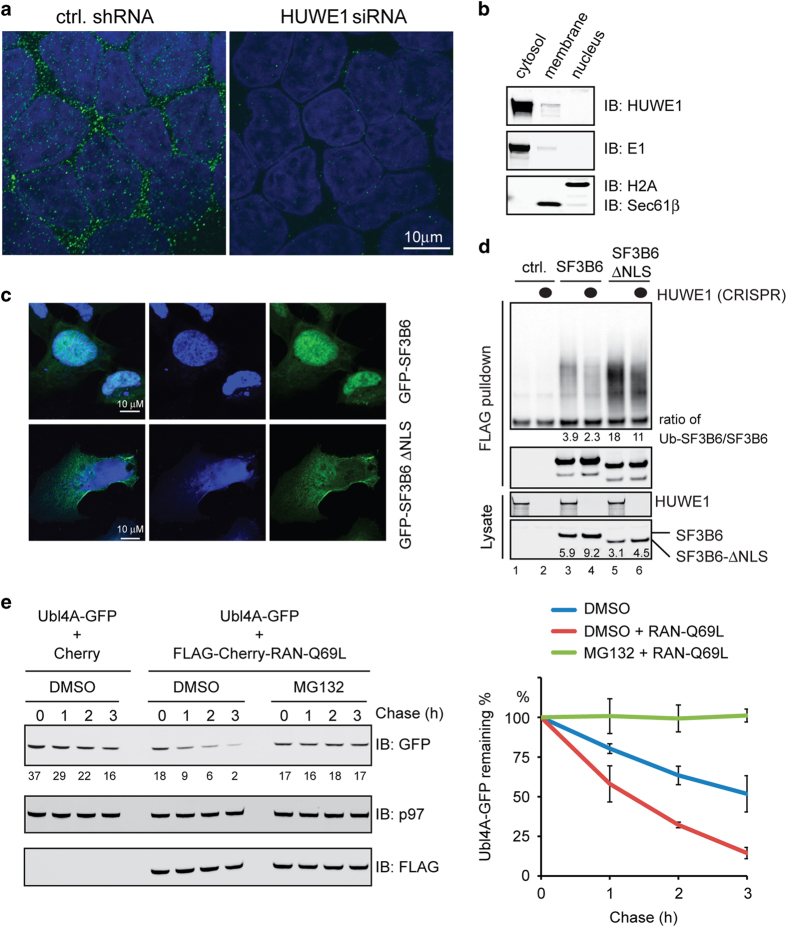
References
-
- Hegde RS, Lingappa VR. Membrane protein biogenesis: regulated complexity at the endoplasmic reticulum. Cell 1997; 91: 575–582. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
