

Systemic depletion of L-cyst(e)ine with cyst(e)inase increases reactive oxygen species and suppresses tumor growth
- PMID: 27869804
- PMCID: PMC5218918
- DOI: 10.1038/nm.4232
Systemic depletion of L-cyst(e)ine with cyst(e)inase increases reactive oxygen species and suppresses tumor growth
Abstract
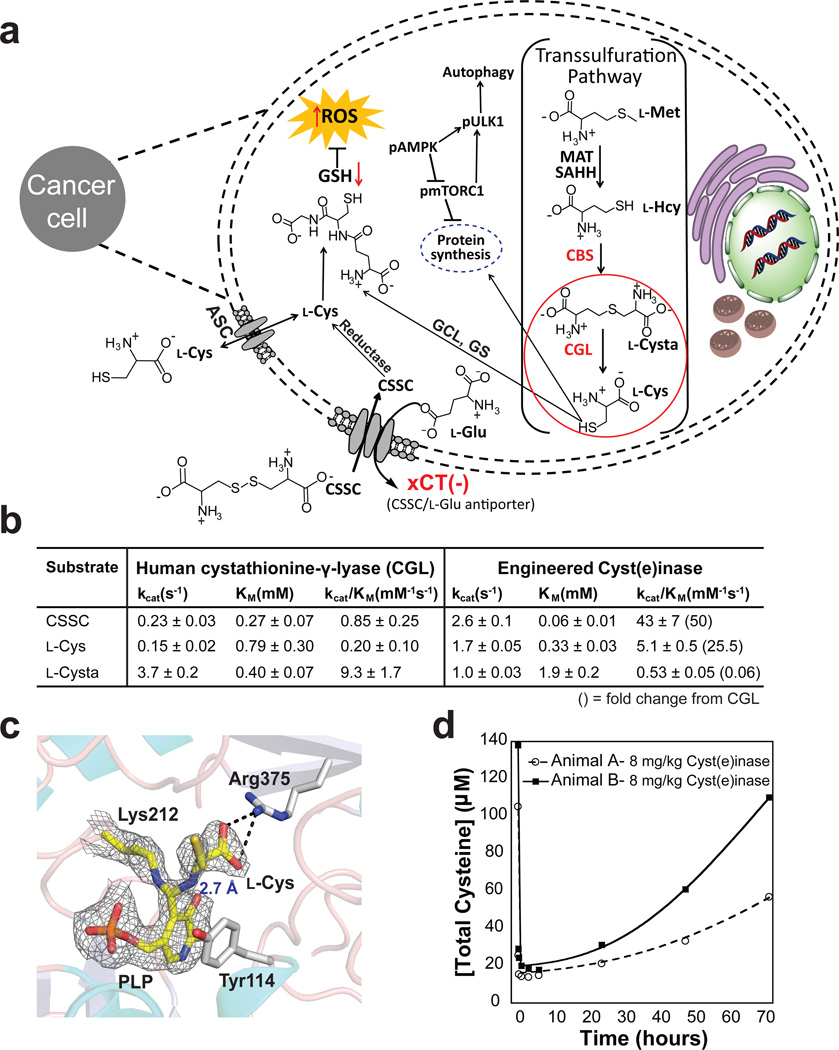
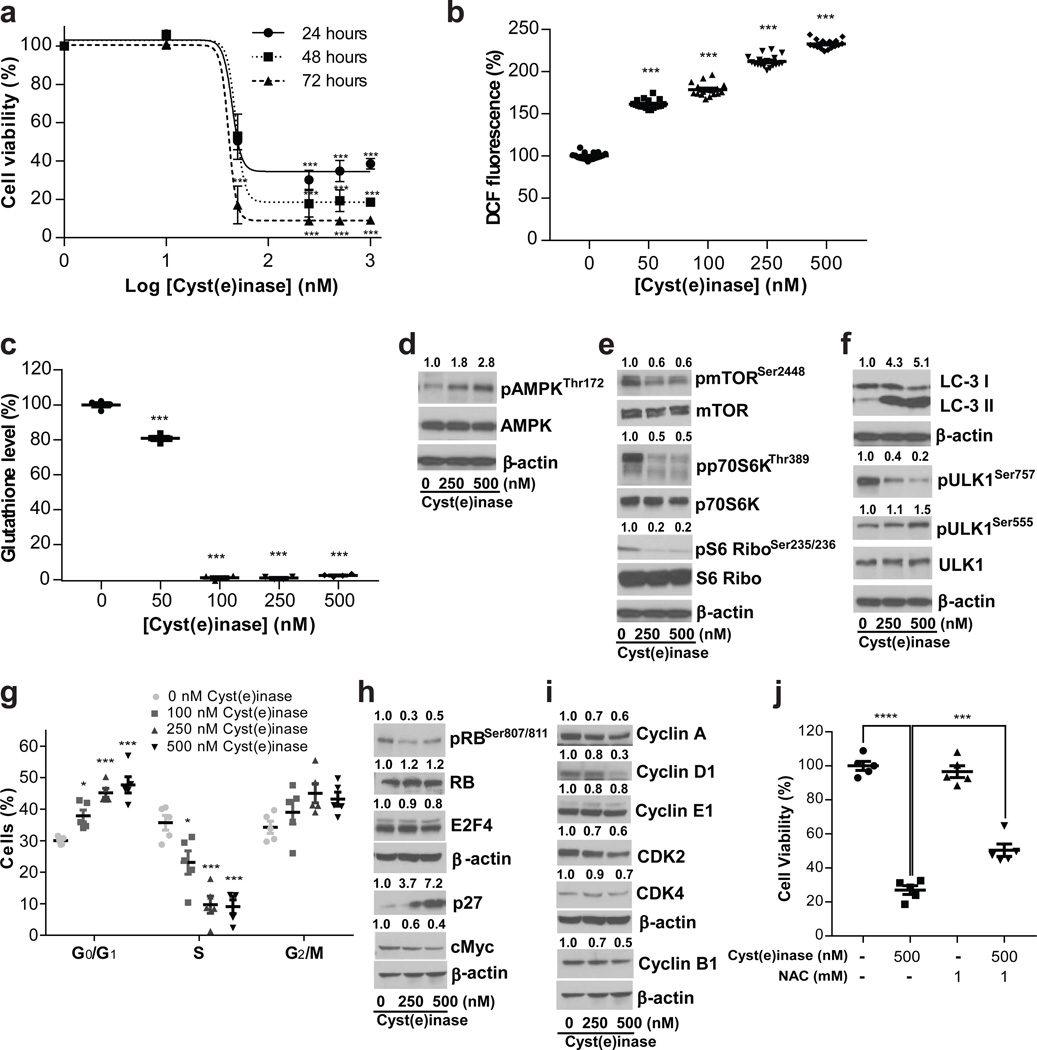
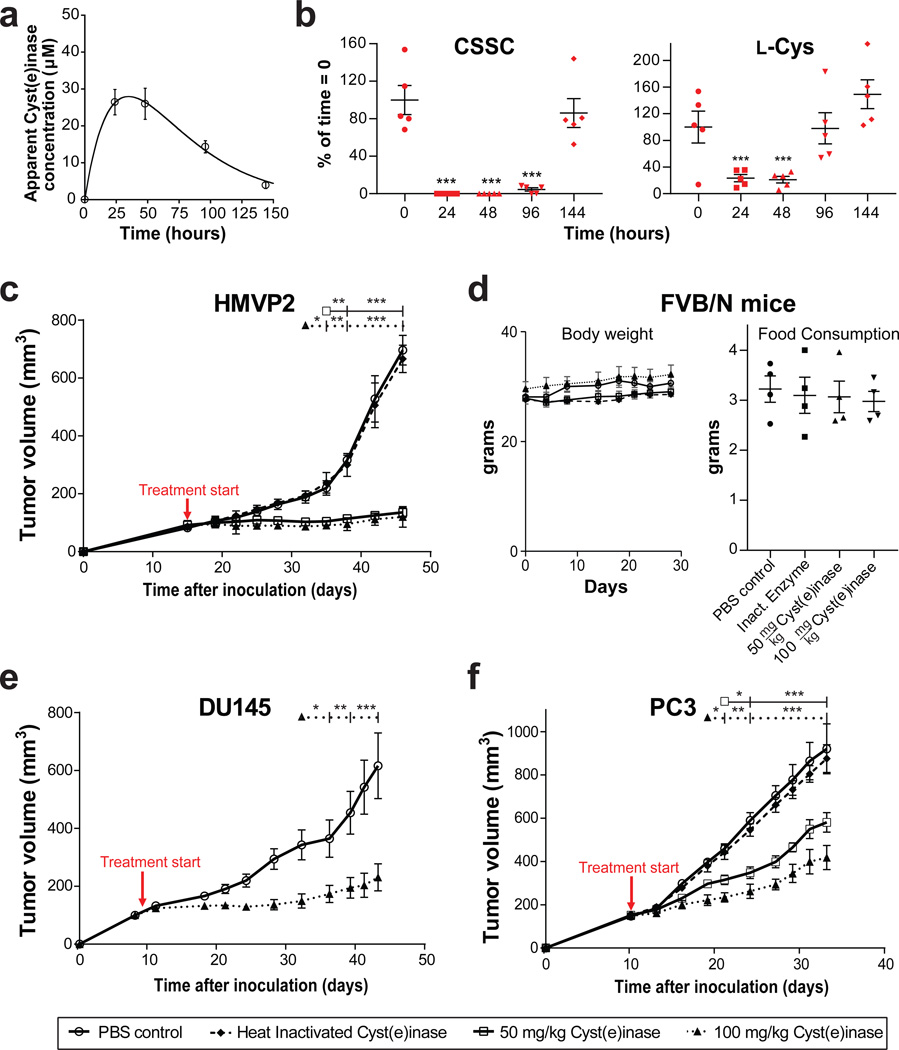
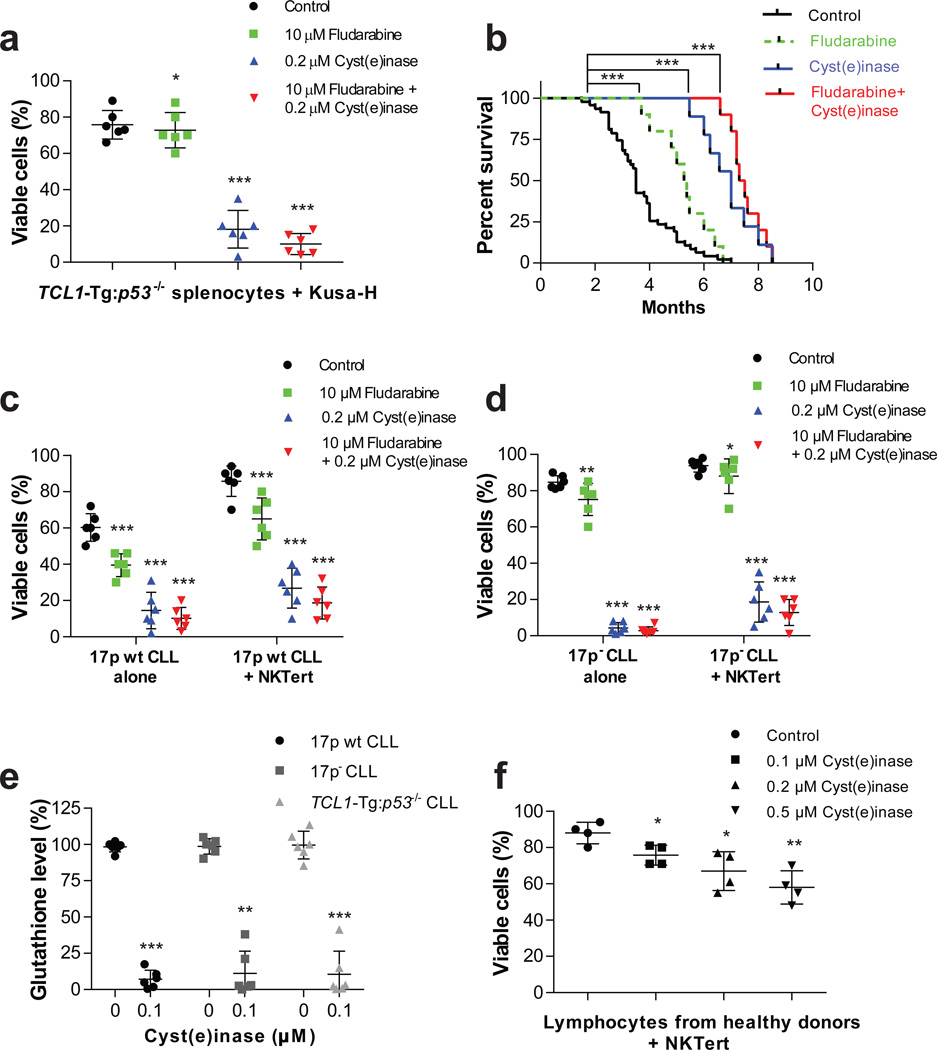
Cancer cells experience higher oxidative stress from reactive oxygen species (ROS) than do non-malignant cells because of genetic alterations and abnormal growth; as a result, maintenance of the antioxidant glutathione (GSH) is essential for their survival and proliferation. Under conditions of elevated ROS, endogenous L-cysteine (L-Cys) production is insufficient for GSH synthesis. This necessitates uptake of L-Cys that is predominantly in its disulfide form, L-cystine (CSSC), via the xCT(-) transporter. We show that administration of an engineered and pharmacologically optimized human cyst(e)inase enzyme mediates sustained depletion of the extracellular L-Cys and CSSC pool in mice and non-human primates. Treatment with this enzyme selectively causes cell cycle arrest and death in cancer cells due to depletion of intracellular GSH and ensuing elevated ROS; yet this treatment results in no apparent toxicities in mice even after months of continuous treatment. Cyst(e)inase suppressed the growth of prostate carcinoma allografts, reduced tumor growth in both prostate and breast cancer xenografts and doubled the median survival time of TCL1-Tg:p53-/- mice, which develop disease resembling human chronic lymphocytic leukemia. It was observed that enzyme-mediated depletion of the serum L-Cys and CSSC pool suppresses the growth of multiple tumors, yet is very well tolerated for prolonged periods, suggesting that cyst(e)inase represents a safe and effective therapeutic modality for inactivating antioxidant cellular responses in a wide range of malignancies.
Figures
References
-
- Trachootham D, Alexandre J, Huang P. Targeting cancer cells by ROS-mediated mechanisms: a radical therapeutic approach? Nature Reviews Drug Discovery. 2009;8:579–591. - PubMed
-
- Dixon SJ, Stockwell BR. The role of iron and reactive oxygen species in cell death. Nature chemical biology. 2014;10:9–17. - PubMed
-
- Harris IS, et al. Glutathione and thioredoxin antioxidant pathways synergize to drive cancer initiation and progression. Cancer cell. 2015;27:211–222. - PubMed
-
- Liu H, Zhi Y, Geng G, Yu Z, Xu H. Effect of phenethyl isothiocyanate given at different duration of gestation on the outcome of pregnancy in rats. Journal of hygiene research. 2011;40:283–286. - PubMed
-
- Reliene R, Schiestl RH. Glutathione depletion by buthionine sulfoximine induces DNA deletions in mice. Carcinogenesis. 2006;27:240–244. - PubMed
Online Methods References
-
- Minor W, Otwinowski Z. Methods in Enzymology, Macromolecular Crystallography. New York: Academic Press; 1997. HKL2000 (Denzo-SMN) Software Package. Processing of X-ray Diffraction Data Collected in Oscillation Mode. - PubMed
-
- DeLano WL. The PyMOL molecular graphics system. 2002
-
- Cheson BD, et al. Guidelines for clinical protocols for chronic lymphocytic leukemia: Recommendations of the national cancer institute-sponsored working group. American journal of hematology. 1988;29:152–163. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
