

The genomic basis of circadian and circalunar timing adaptations in a midge
- PMID: 27871090
- PMCID: PMC5133387
- DOI: 10.1038/nature20151
The genomic basis of circadian and circalunar timing adaptations in a midge
Abstract
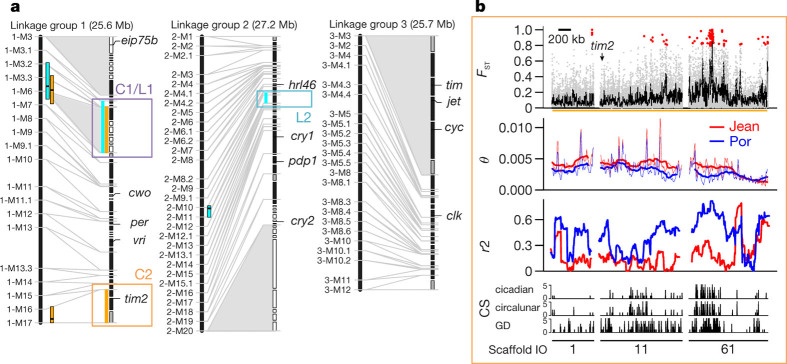
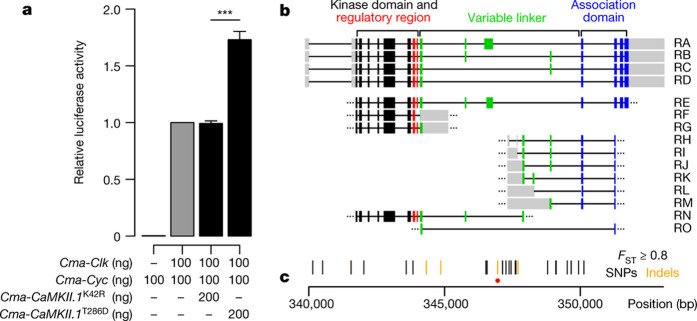
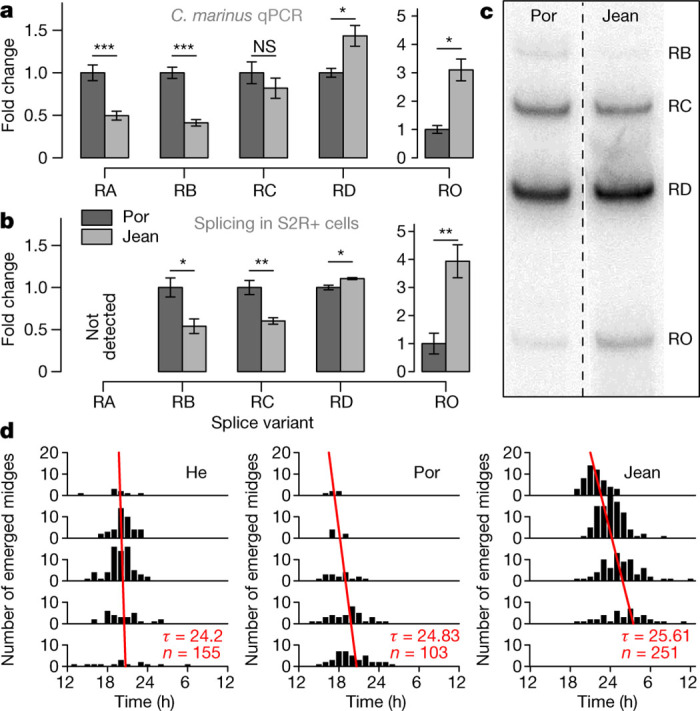
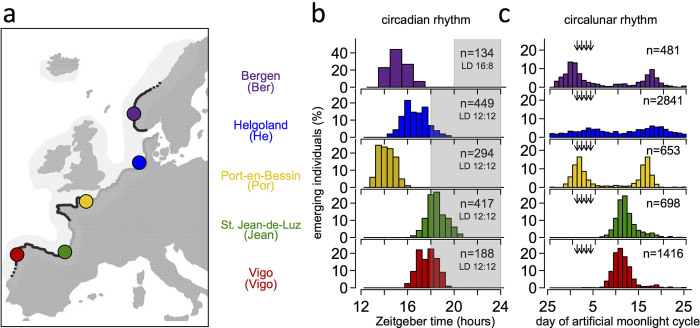
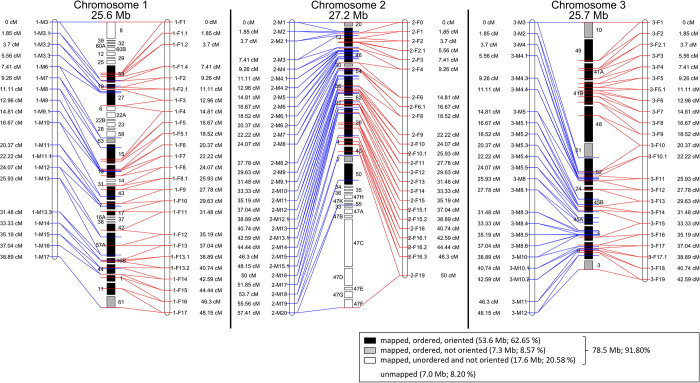
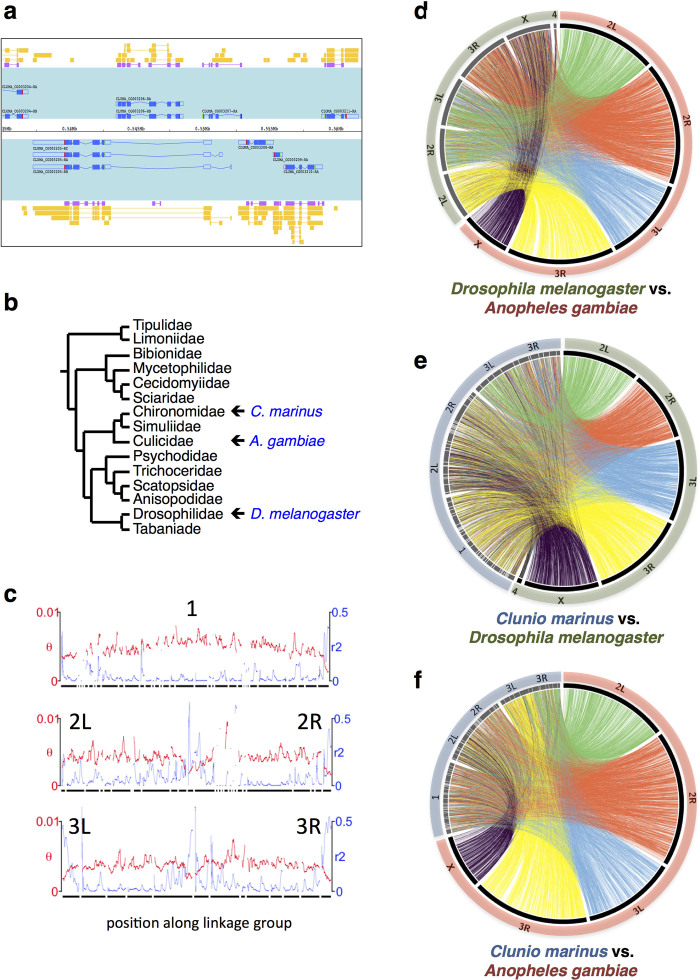
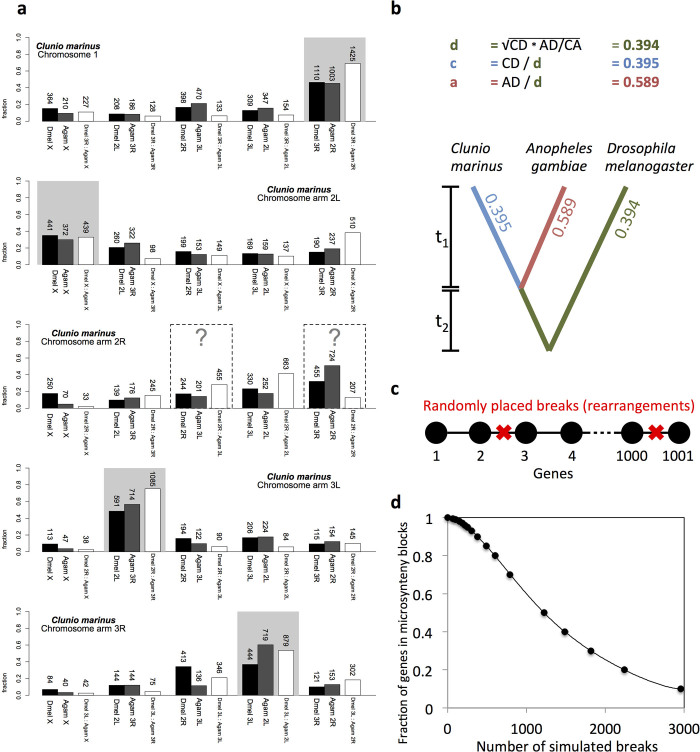
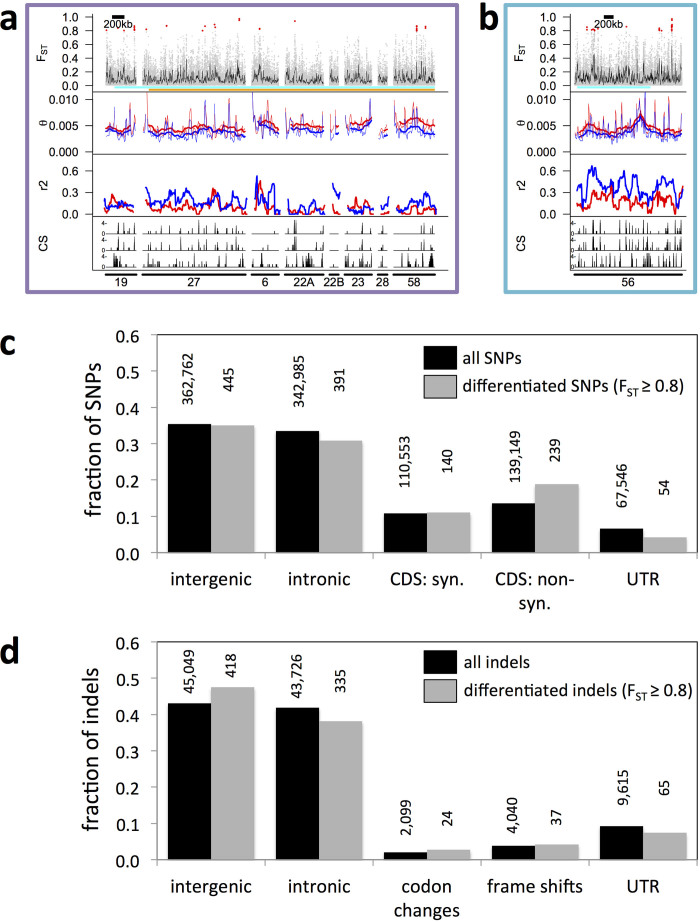
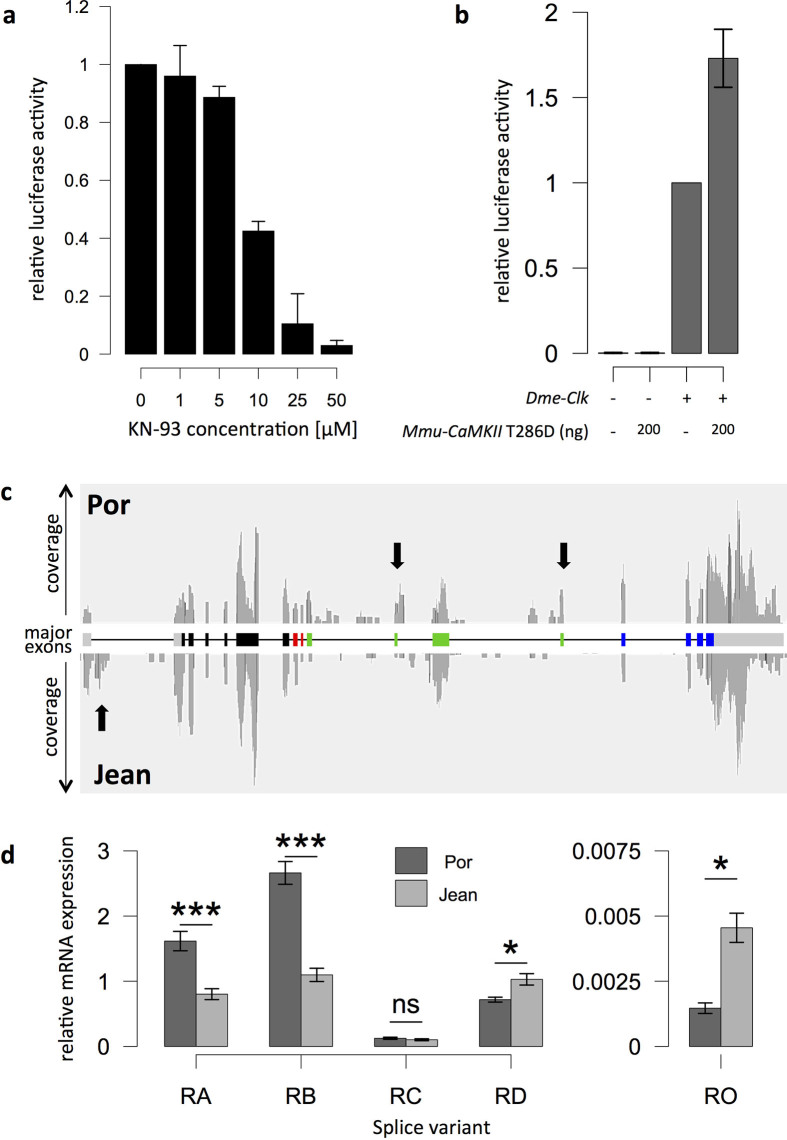
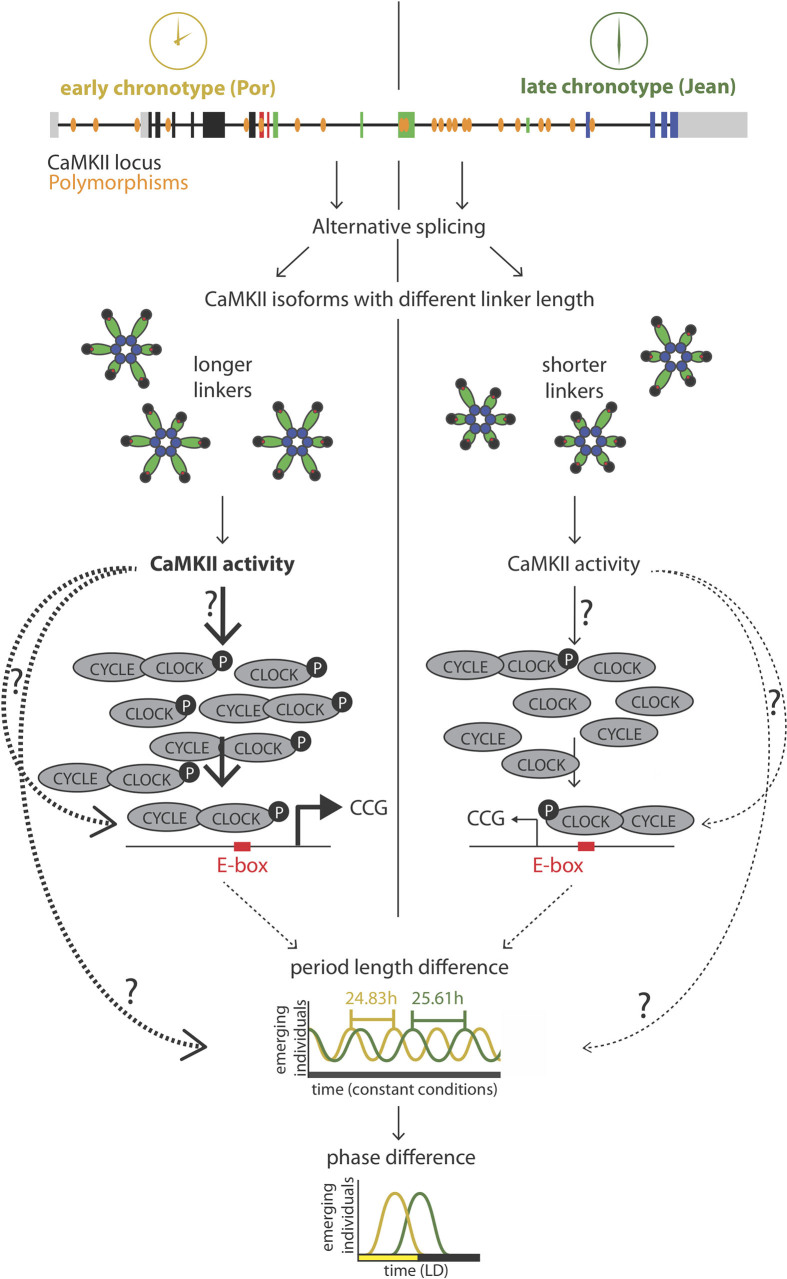
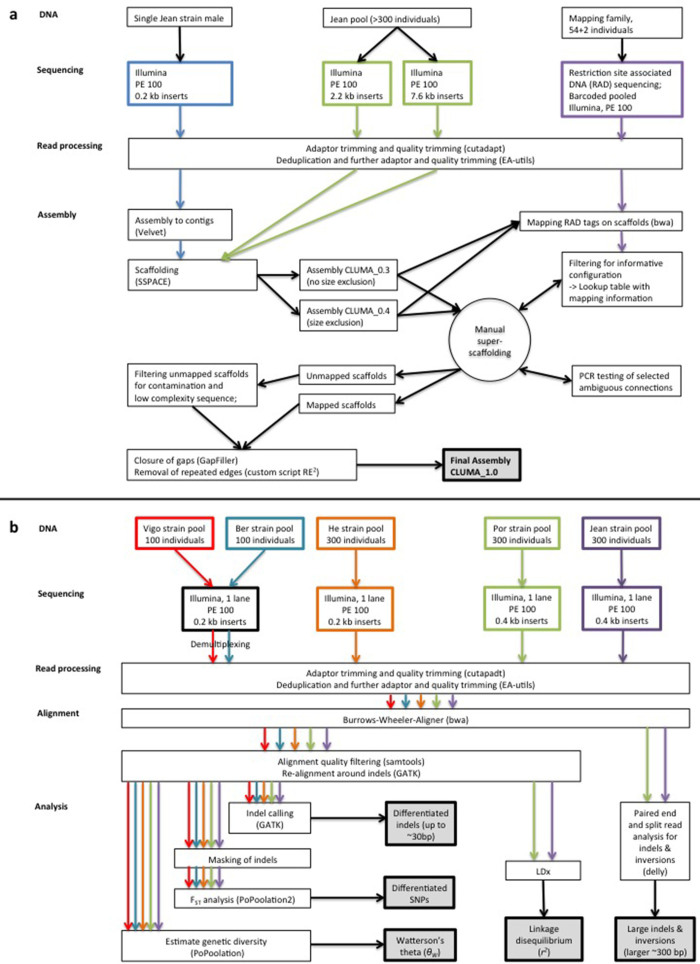
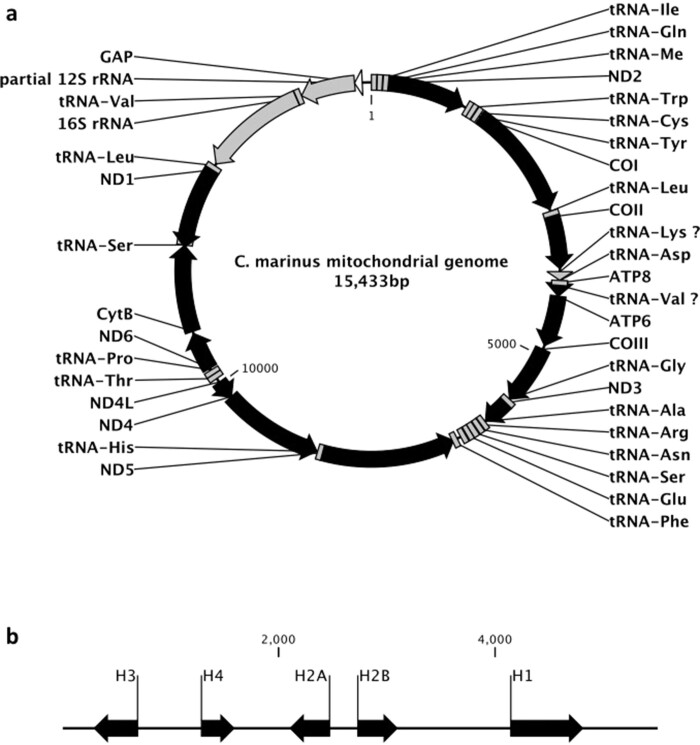
Organisms use endogenous clocks to anticipate regular environmental cycles, such as days and tides. Natural variants resulting in differently timed behaviour or physiology, known as chronotypes in humans, have not been well characterized at the molecular level. We sequenced the genome of Clunio marinus, a marine midge whose reproduction is timed by circadian and circalunar clocks. Midges from different locations show strain-specific genetic timing adaptations. We examined genetic variation in five C. marinus strains from different locations and mapped quantitative trait loci for circalunar and circadian chronotypes. The region most strongly associated with circadian chronotypes generates strain-specific differences in the abundance of calcium/calmodulin-dependent kinase II.1 (CaMKII.1) splice variants. As equivalent variants were shown to alter CaMKII activity in Drosophila melanogaster, and C. marinus (Cma)-CaMKII.1 increases the transcriptional activity of the dimer of the circadian proteins Cma-CLOCK and Cma-CYCLE, we suggest that modulation of alternative splicing is a mechanism for natural adaptation in circadian timing.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

References
-
- Neumann D. Die lunare und tägliche Schlüpfperiodik der Mücke Clunio. Steuerung und Abstimmung auf die Gezeitenperiodik. Z. Vgl. Physiol. 1966;53:1–61.
-
- Neumann D. Temperature compensation of circasemilunar timing in the intertidal insect Clunio. J. Comp. Physiol. 1988;163:671–676.
-
- ADMIRALTY Tide Tables (UKHO, 2014)
-
- Neumann D. Genetic adaptation in emergence time of Clunio populations to different tidal conditions. Helgoländer Wiss. Meeresunters. 1967;15:163–171.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
