

The Transcription Factor ATHB5 Affects GA-Mediated Plasticity in Hypocotyl Cell Growth during Seed Germination
- PMID: 27872245
- PMCID: PMC5210717
- DOI: 10.1104/pp.16.01099
The Transcription Factor ATHB5 Affects GA-Mediated Plasticity in Hypocotyl Cell Growth during Seed Germination
Abstract
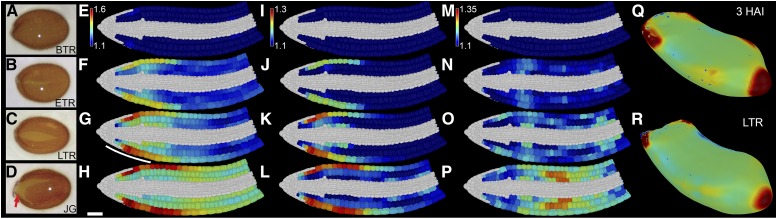
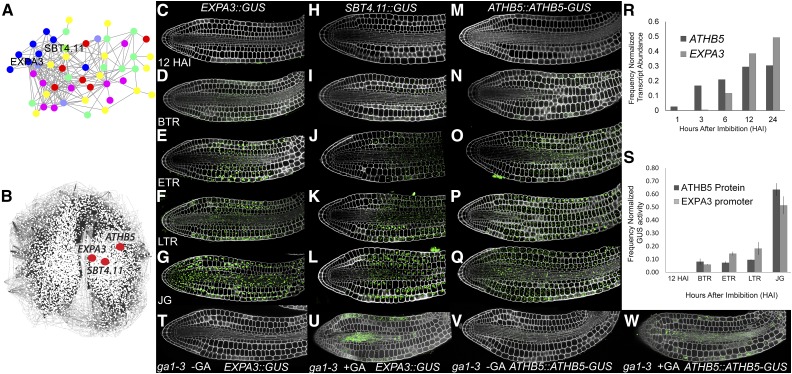
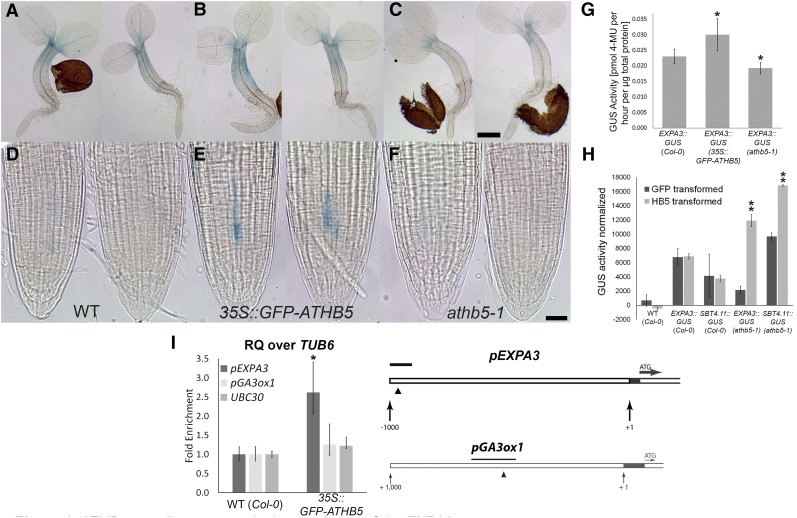
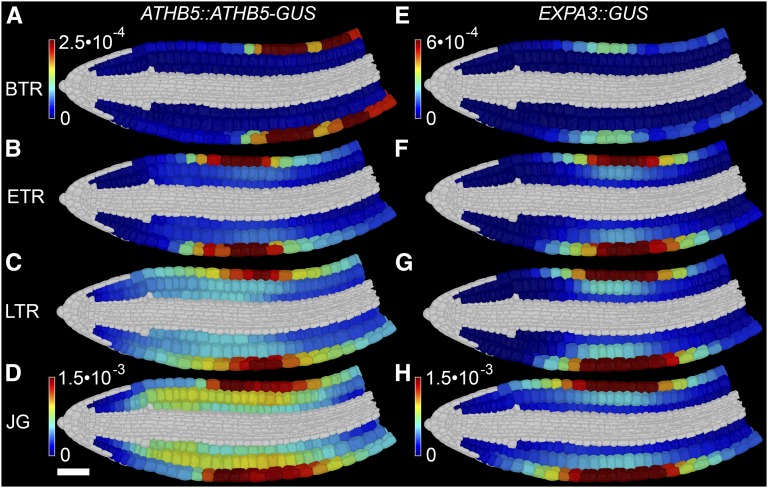
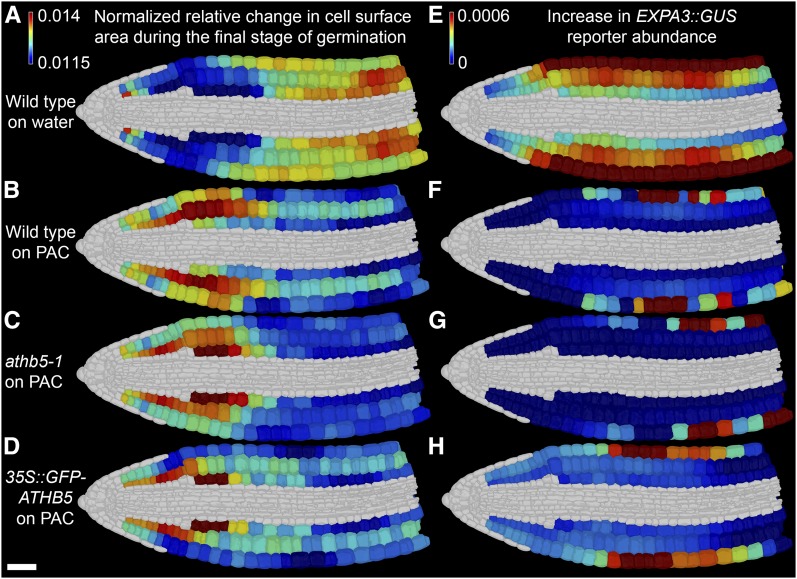
Gibberellic acid (GA)-mediated cell expansion initiates the seed-to-seedling transition in plants and is repressed by DELLA proteins. Using digital single-cell analysis, we identified a cellular subdomain within the midhypocotyl, whose expansion drives the final step of this developmental transition under optimal conditions. Using network inference, the transcription factor ATHB5 was identified as a genetic factor whose localized expression promotes GA-mediated expansion specifically within these cells. Both this protein and its putative growth-promoting target EXPANSIN3 are repressed by DELLA, and coregulated at single-cell resolution during seed germination. The cellular domains of hormone sensitivity were explored within the Arabidopsis (Arabidopsis thaliana) embryo by putting seeds under GA-limiting conditions and quantifying cellular growth responses. The middle and upper hypocotyl have a greater requirement for GA to promote cell expansion than the lower embryo axis. Under these conditions, germination was still completed following enhanced growth within the radicle and lower axis. Under GA-limiting conditions, the athb5 mutant did not show a phenotype at the level of seed germination, but it did at a cellular level with reduced cell expansion in the hypocotyl relative to the wild type. These data reveal that the spatiotemporal cell expansion events driving this transition are not determinate, and the conditional use of GA-ATHB5-mediated hypocotyl growth under optimal conditions may be used to optionally support rapid seedling growth. This study demonstrates that multiple genetic and spatiotemporal cell expansion mechanisms underlie the seed to seedling transition in Arabidopsis.
© 2017 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Achard P, Cheng H, De Grauwe L, Decat J, Schoutteten H, Moritz T, Van Der Straeten D, Peng J, Harberd NP (2006) Integration of plant responses to environmentally activated phytohormonal signals. Science 311: 91–94 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BB/N009754/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/G015066/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/J017604/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/M01116X/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/L010232/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
