

REV-ERBα regulates Fgf21 expression in the liver via hepatic nuclear factor 6
- PMID: 27875243
- PMCID: PMC5278426
- DOI: 10.1242/bio.021519
REV-ERBα regulates Fgf21 expression in the liver via hepatic nuclear factor 6
Abstract
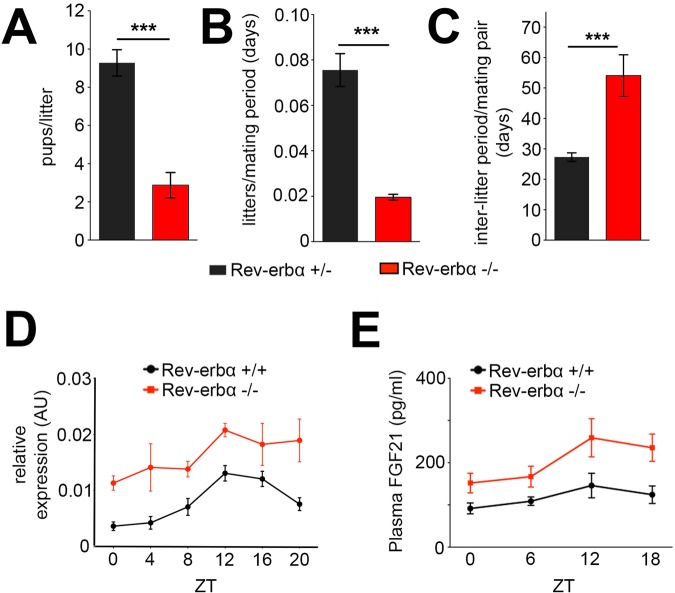
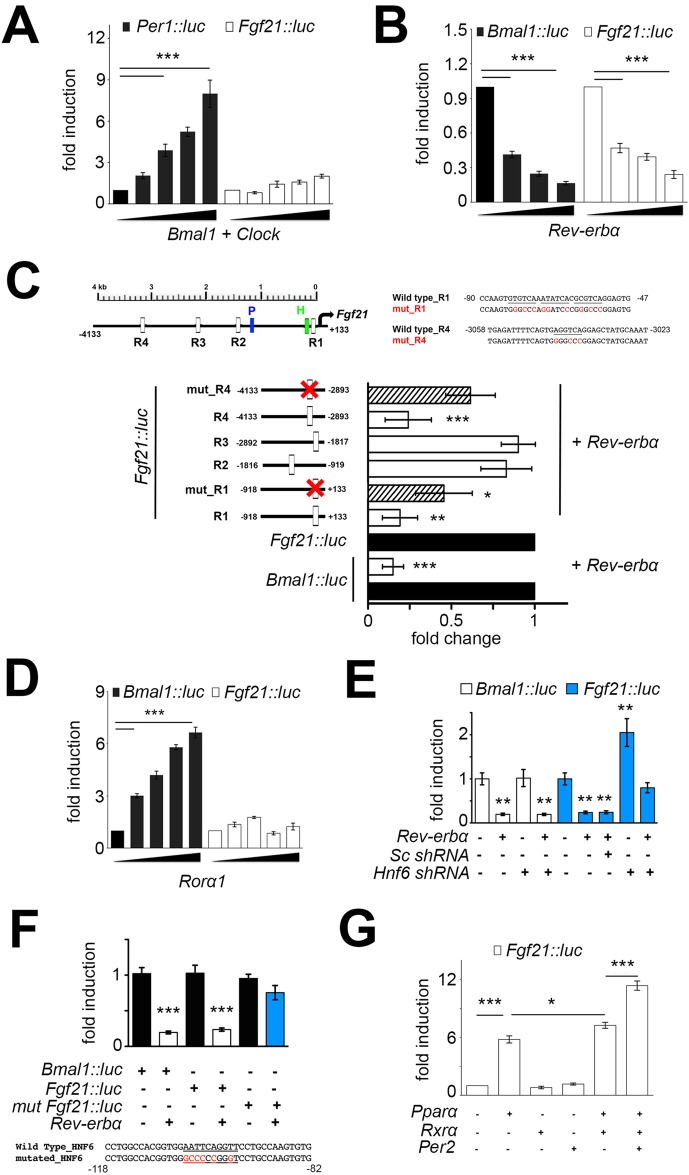
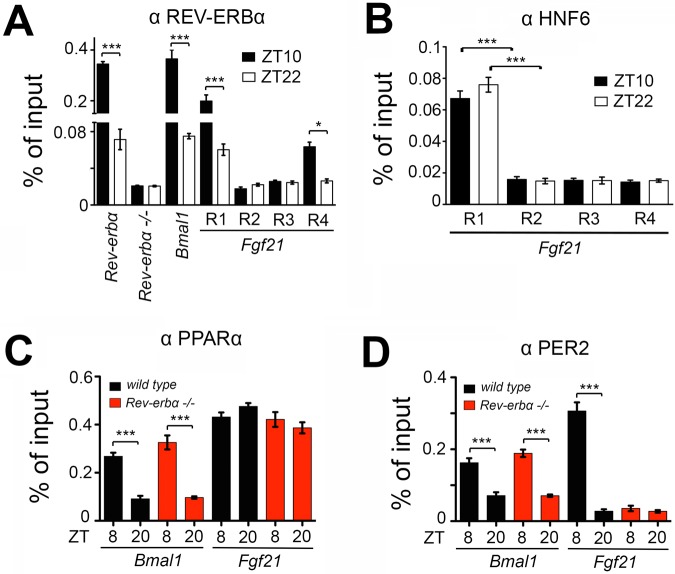
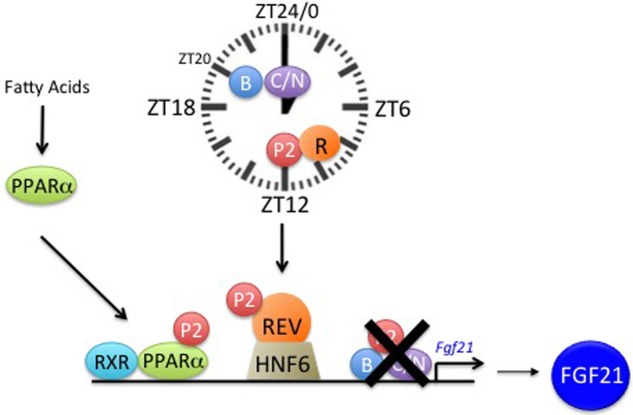
The circadian clock contributes to the timing of many body functions including metabolism and reproduction. The hepatokine fibroblast growth factor 21 (FGF21) is a critical metabolic regulator involved in modulation of fertility. Here we show that lack of the clock component REV-ERBα elevates FGF21 levels in liver and plasma. At the molecular level, REV-ERBα modulates the expression of FGF21 via the liver-specific hepatic nuclear factor 6 (HNF6). We conclude that REV-ERBα regulates metabolism and reproduction, at least in part, via regulation of Fgf21.
Keywords: Circadian clock; Physiology; Transcription.
© 2017. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
References
-
- Canaple L., Rambaud J., Dkhissi-Benyahya O., Rayet B., Tan N. S., Michalik L., Delaunay F., Wahli W. and Laudet V. (2006). Reciprocal regulation of brain and muscle Arnt-like protein 1 and peroxisome proliferator-activated receptor alpha defines a novel positive feedback loop in the rodent liver circadian clock. Mol. Endocrinol. 20, 1715-1727. 10.1210/me.2006-0052 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
