

Single Binding Mode Integration of Hemicellulose-degrading Enzymes via Adaptor Scaffoldins in Ruminococcus flavefaciens Cellulosome
- PMID: 27875311
- PMCID: PMC5207176
- DOI: 10.1074/jbc.M116.761643
Single Binding Mode Integration of Hemicellulose-degrading Enzymes via Adaptor Scaffoldins in Ruminococcus flavefaciens Cellulosome
Abstract
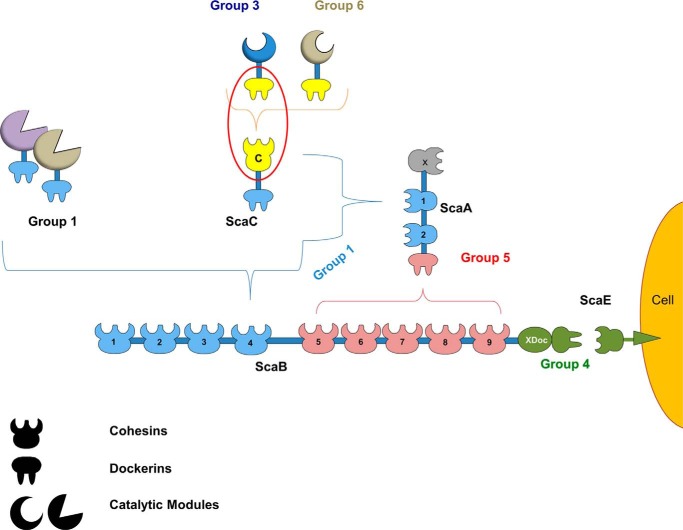
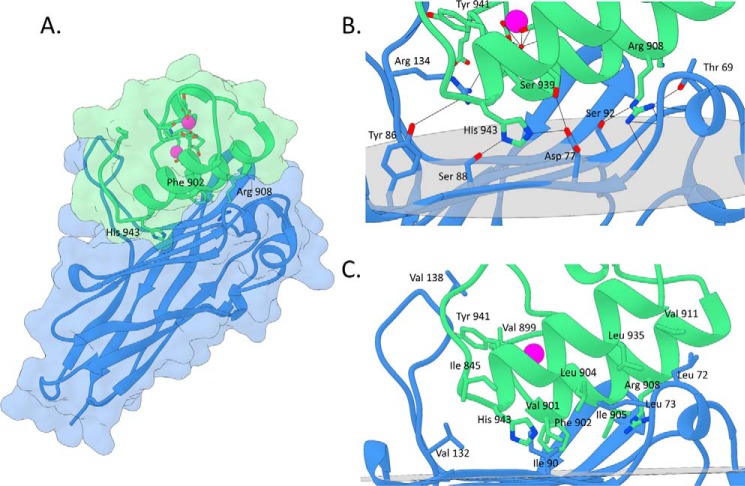
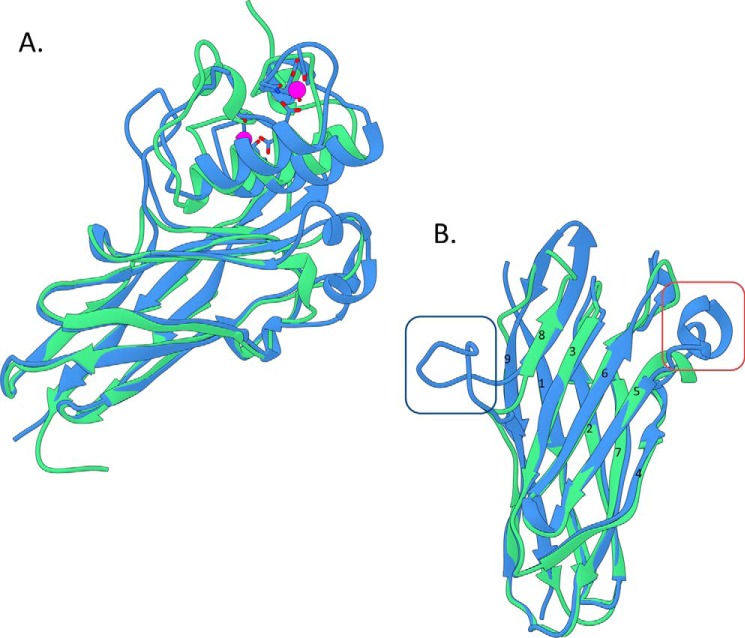
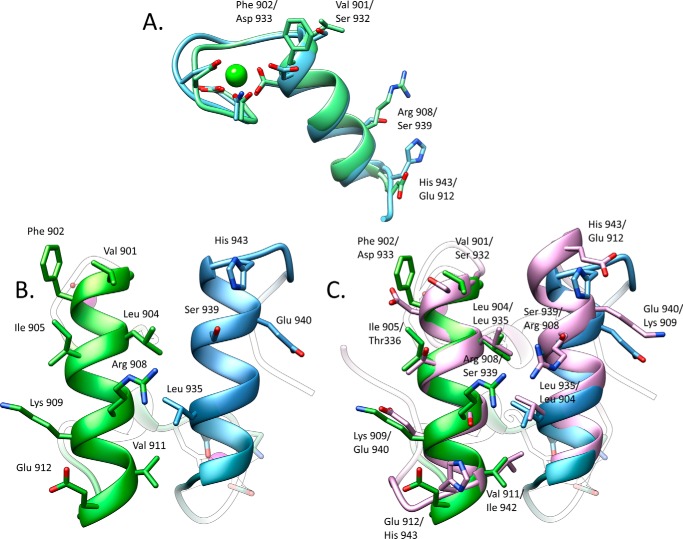
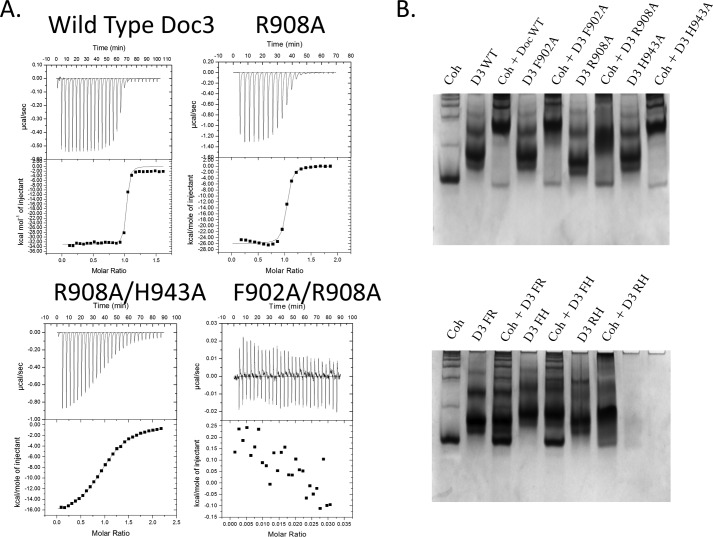
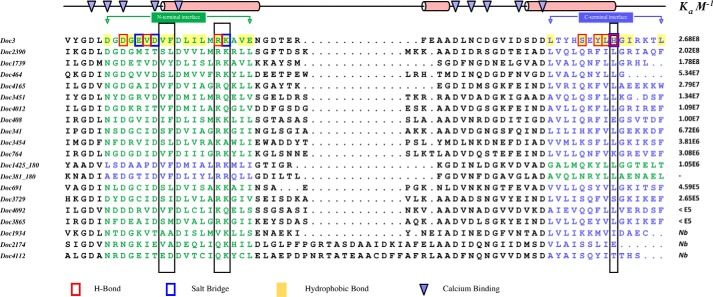
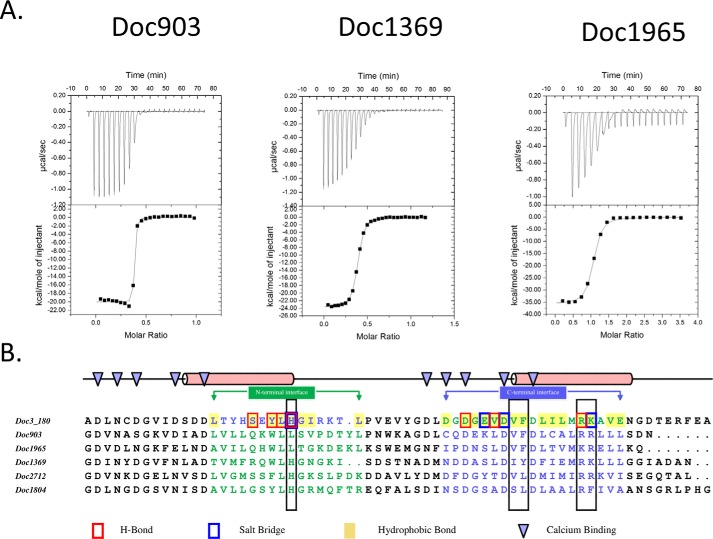
The assembly of one of Nature's most elaborate multienzyme complexes, the cellulosome, results from the binding of enzyme-borne dockerins to reiterated cohesin domains located in a non-catalytic primary scaffoldin. Generally, dockerins present two similar cohesin-binding interfaces that support a dual binding mode. The dynamic integration of enzymes in cellulosomes, afforded by the dual binding mode, is believed to incorporate additional flexibility in highly populated multienzyme complexes. Ruminococcus flavefaciens, the primary degrader of plant structural carbohydrates in the rumen of mammals, uses a portfolio of more than 220 different dockerins to assemble the most intricate cellulosome known to date. A sequence-based analysis organized R. flavefaciens dockerins into six groups. Strikingly, a subset of R. flavefaciens cellulosomal enzymes, comprising dockerins of groups 3 and 6, were shown to be indirectly incorporated into primary scaffoldins via an adaptor scaffoldin termed ScaC. Here, we report the crystal structure of a group 3 R. flavefaciens dockerin, Doc3, in complex with ScaC cohesin. Doc3 is unusual as it presents a large cohesin-interacting surface that lacks the structural symmetry required to support a dual binding mode. In addition, dockerins of groups 3 and 6, which bind exclusively to ScaC cohesin, display a conserved mechanism of protein recognition that is similar to Doc3. Groups 3 and 6 dockerins are predominantly appended to hemicellulose-degrading enzymes. Thus, single binding mode dockerins interacting with adaptor scaffoldins exemplify an evolutionary pathway developed by R. flavefaciens to recruit hemicellulases to the sophisticated cellulosomes acting in the gastrointestinal tract of mammals.
Keywords: cellulase; cellulose; cellulosome; cohesin; dockerin; protein structure; protein-protein interaction.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Bayer E. A., Lamed R., and Himmel M. E. (2007) The potential of cellulases and cellulosomes for cellulosic waste management. Curr. Opin. Biotechnol. 18, 237–245 - PubMed
-
- Fontes C. M., and Gilbert H. J. (2010) Cellulosomes: highly efficient nanomachines designed to deconstruct plant cell wall complex carbohydrates. Annu. Rev. Biochem. 79, 655–681 - PubMed
-
- Bayer E. A., Belaich J.-P., Shoham Y., and Lamed R. (2004) The cellulosomes: multienzyme machines for degradation of plant cell wall polysaccharides. Annu. Rev. Microbiol. 58, 521–554 - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
