

Palmitoylated APP Forms Dimers, Cleaved by BACE1
- PMID: 27875558
- PMCID: PMC5119739
- DOI: 10.1371/journal.pone.0166400
Palmitoylated APP Forms Dimers, Cleaved by BACE1
Erratum in
-
Correction: Palmitoylated APP Forms Dimers, Cleaved by BACE1.PLoS One. 2024 Feb 29;19(2):e0299972. doi: 10.1371/journal.pone.0299972. eCollection 2024. PLoS One. 2024. PMID: 38421989 Free PMC article.
Abstract
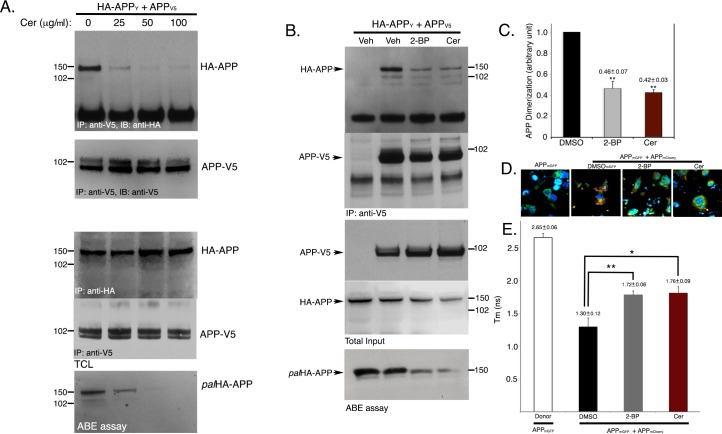
A major rate-limiting step for Aβ generation and deposition in Alzheimer's disease brains is BACE1-mediated cleavage (β-cleavage) of the amyloid precursor protein (APP). We previously reported that APP undergoes palmitoylation at two cysteine residues (Cys186 and Cys187) in the E1-ectodomain. 8-10% of total APP is palmitoylated in vitro and in vivo. Palmitoylated APP (palAPP) shows greater preference for β-cleavage than total APP in detergent resistant lipid rafts. Protein palmitoylation is known to promote protein dimerization. Since dimerization of APP at its E1-ectodomain results in elevated BACE1-mediated cleavage of APP, we have now investigated whether palmitoylation of APP affects its dimerization and whether this leads to elevated β-cleavage of the protein. Here we report that over 90% of palAPP is dimerized while only ~20% of total APP forms dimers. PalAPP-dimers are predominantly cis-oriented while total APP dimerizes in both cis- and trans-orientation. PalAPP forms dimers 4.5-times more efficiently than total APP. Overexpression of the palmitoylating enzymes DHHC7 and DHHC21 that increase palAPP levels and Aβ release, also increased APP dimerization in cells. Conversely, inhibition of APP palmitoylation by pharmacological inhibitors reduced APP-dimerization in coimmunoprecipitation and FLIM/FRET assays. Finally, in vitro BACE1-activity assays demonstrate that palmitoylation-dependent dimerization of APP promotes β-cleavage of APP in lipid-rich detergent resistant cell membranes (DRMs), when compared to total APP. Most importantly, generation of sAPPβ-sAPPβ dimers is dependent on APP-palmitoylation while total sAPPβ generation is not. Since BACE1 shows preference for palAPP dimers over total APP, palAPP dimers may serve as novel targets for effective β-cleavage inhibitors of APP as opposed to BACE1 inhibitors.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
