

Unlocking sperm chromatin at fertilization requires a dedicated egg thioredoxin in Drosophila
- PMID: 27876811
- PMCID: PMC5122968
- DOI: 10.1038/ncomms13539
Unlocking sperm chromatin at fertilization requires a dedicated egg thioredoxin in Drosophila
Abstract
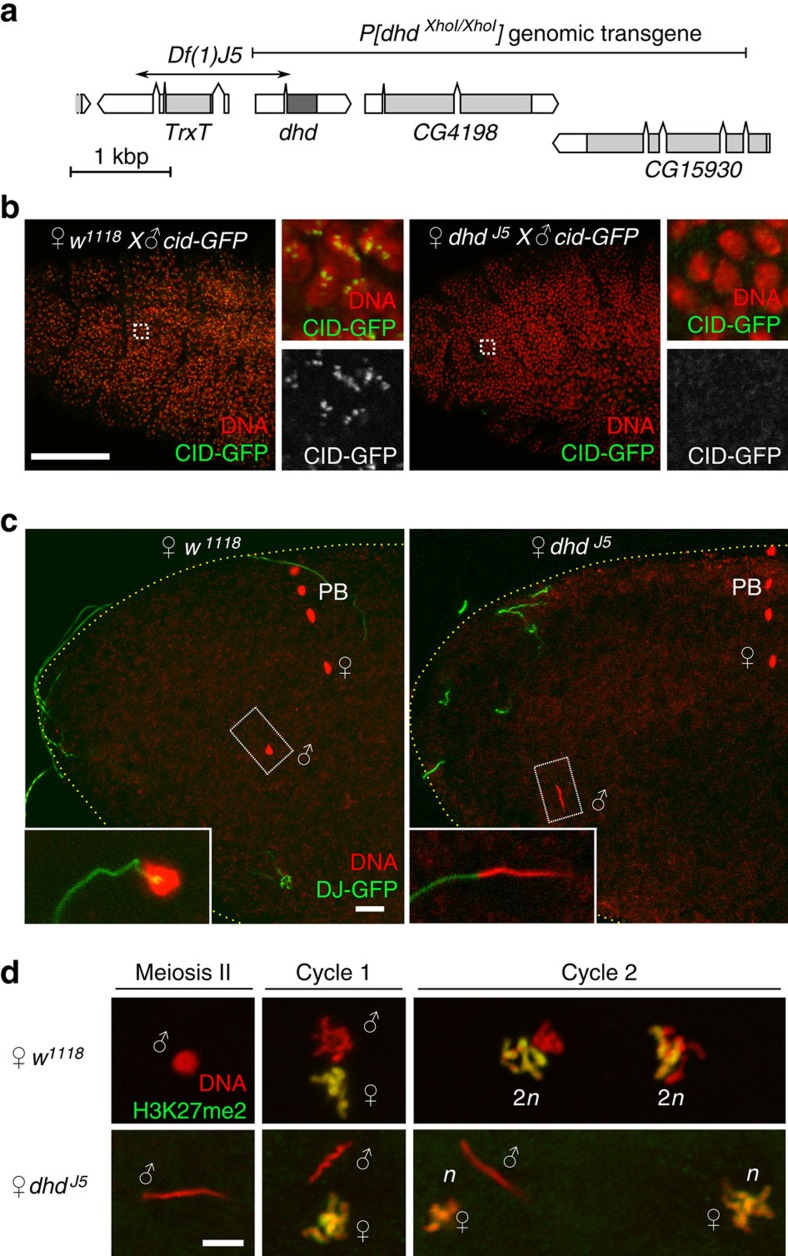
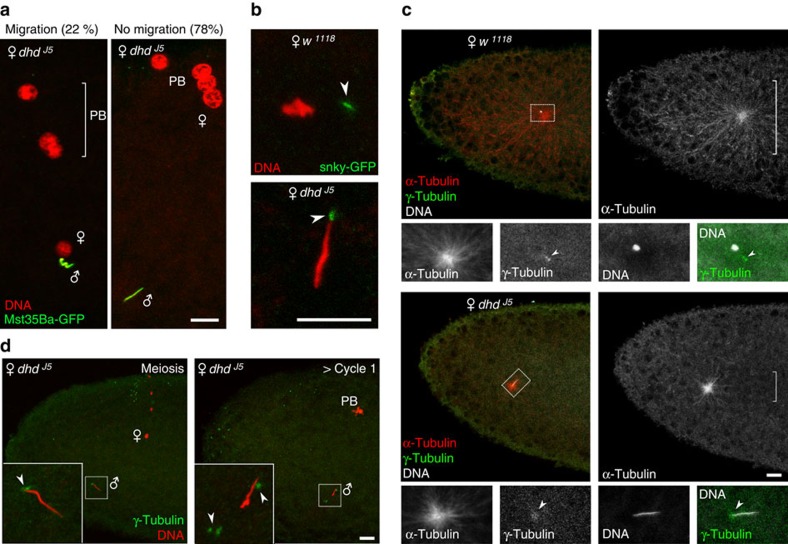
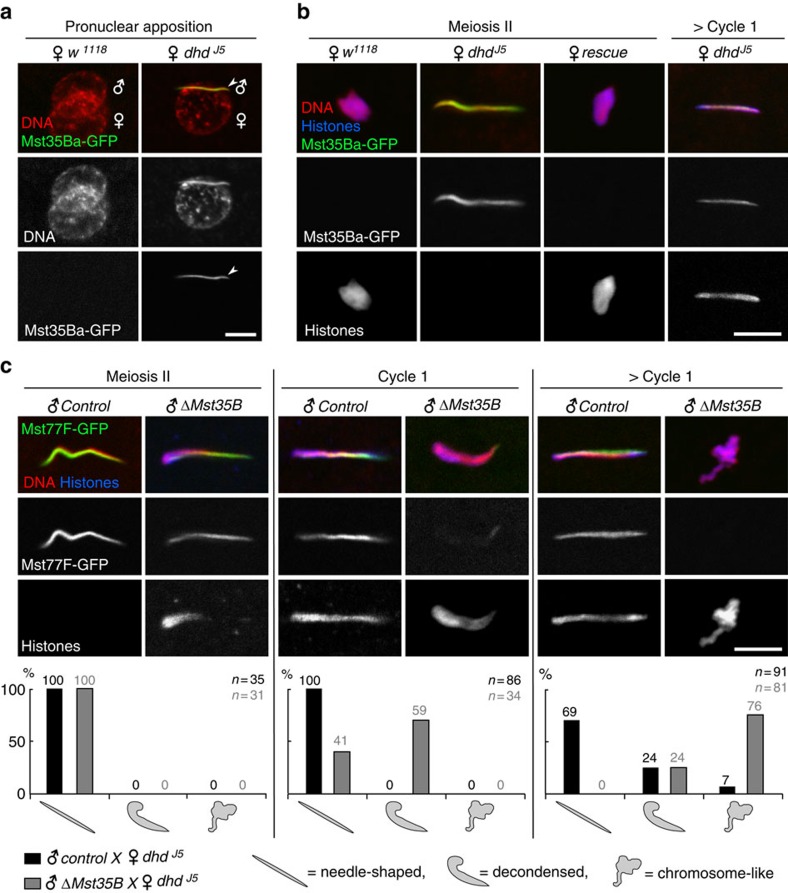
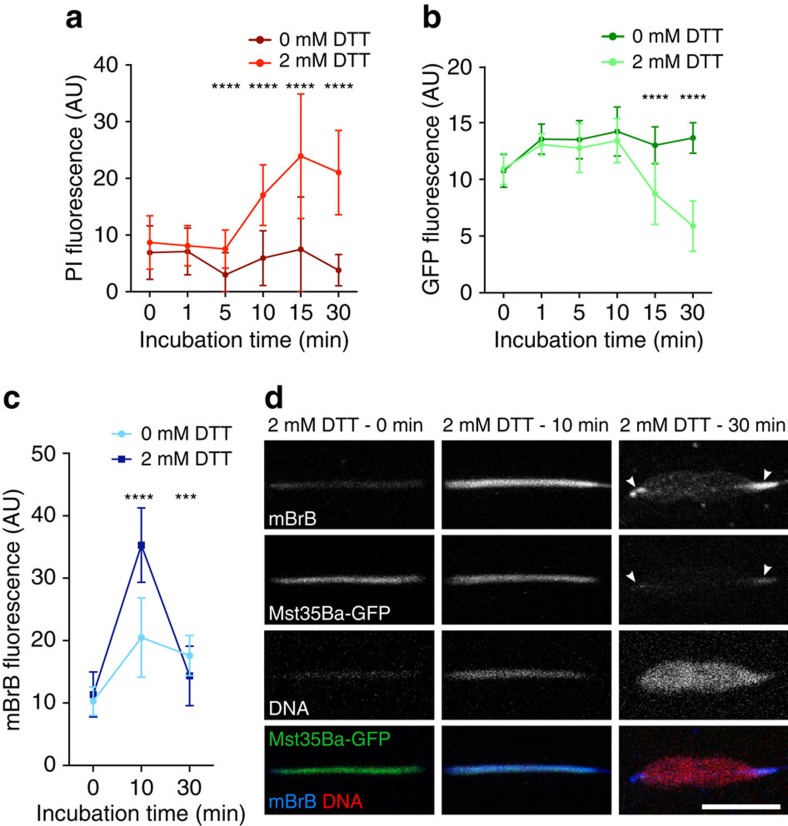
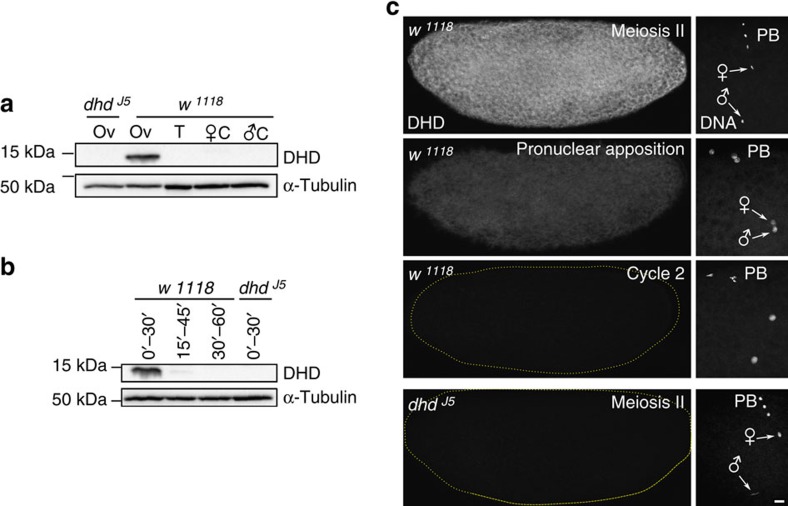
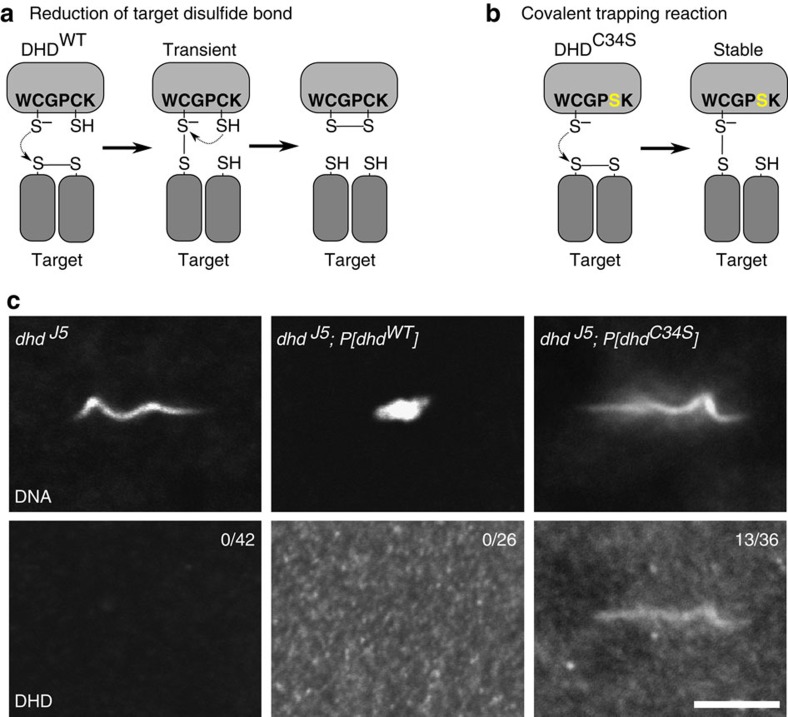
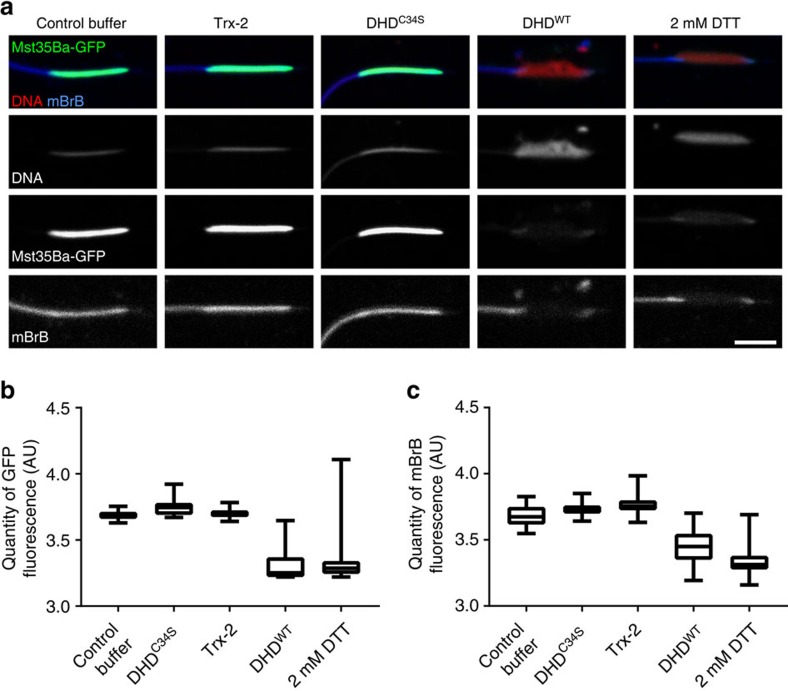
In most animals, the extreme compaction of sperm DNA is achieved after the massive replacement of histones with sperm nuclear basic proteins (SNBPs), such as protamines. In some species, the ultracompact sperm chromatin is stabilized by a network of disulfide bonds connecting cysteine residues present in SNBPs. Studies in mammals have established that the reduction of these disulfide crosslinks at fertilization is required for sperm nuclear decondensation and the formation of the male pronucleus. Here, we show that the Drosophila maternal thioredoxin Deadhead (DHD) is specifically required to unlock sperm chromatin at fertilization. In dhd mutant eggs, the sperm nucleus fails to decondense and the replacement of SNBPs with maternally-provided histones is severely delayed, thus preventing the participation of paternal chromosomes in embryo development. We demonstrate that DHD localizes to the sperm nucleus to reduce its disulfide targets and is then rapidly degraded after fertilization.
Figures
Similar articles
-
Thioredoxin-dependent disulfide bond reduction is required for protamine eviction from sperm chromatin.Genes Dev. 2016 Dec 15;30(24):2651-2656. doi: 10.1101/gad.290916.116. Epub 2016 Dec 28. Genes Dev. 2016. PMID: 28031247 Free PMC article.
-
The Drosophila maternal gene sésame is required for sperm chromatin remodeling at fertilization.Chromosoma. 2001 Nov;110(6):430-40. doi: 10.1007/s004120100161. Epub 2001 Aug 9. Chromosoma. 2001. PMID: 11735001
-
The essential role of Drosophila HIRA for de novo assembly of paternal chromatin at fertilization.PLoS Genet. 2007 Oct;3(10):1991-2006. doi: 10.1371/journal.pgen.0030182. Epub 2007 Sep 10. PLoS Genet. 2007. PMID: 17967064 Free PMC article.
-
The intimate genetics of Drosophila fertilization.Open Biol. 2015 Aug;5(8):150076. doi: 10.1098/rsob.150076. Open Biol. 2015. PMID: 26246493 Free PMC article. Review.
-
Sperm nuclear activation during fertilization.Curr Top Dev Biol. 1999;46:133-78. doi: 10.1016/s0070-2153(08)60328-2. Curr Top Dev Biol. 1999. PMID: 10417879 Review.
Cited by
-
Distinct spermiogenic phenotypes underlie sperm elimination in the Segregation Distorter meiotic drive system.PLoS Genet. 2021 Jul 6;17(7):e1009662. doi: 10.1371/journal.pgen.1009662. eCollection 2021 Jul. PLoS Genet. 2021. PMID: 34228705 Free PMC article.
-
Structures of the germline-specific Deadhead and thioredoxin T proteins from Drosophila melanogaster reveal unique features among thioredoxins.IUCrJ. 2021 Feb 11;8(Pt 2):281-294. doi: 10.1107/S2052252521000221. eCollection 2021 Mar 1. IUCrJ. 2021. PMID: 33708404 Free PMC article.
-
Redox Regulation and Oxidative Stress: The Particular Case of the Stallion Spermatozoa.Antioxidants (Basel). 2019 Nov 19;8(11):567. doi: 10.3390/antiox8110567. Antioxidants (Basel). 2019. PMID: 31752408 Free PMC article. Review.
-
An orphan gene is essential for efficient sperm entry into eggs in Drosophila melanogaster.Genetics. 2025 Mar 17;229(3):iyaf008. doi: 10.1093/genetics/iyaf008. Genetics. 2025. PMID: 39903197
-
The Lid/KDM5 histone demethylase complex activates a critical effector of the oocyte-to-zygote transition.PLoS Genet. 2020 Mar 5;16(3):e1008543. doi: 10.1371/journal.pgen.1008543. eCollection 2020 Mar. PLoS Genet. 2020. PMID: 32134927 Free PMC article.
References
-
- Miller D., Brinkworth M. & Iles D. Paternal DNA packaging in spermatozoa: more than the sum of its parts? DNA, histones, protamines and epigenetics. Reproduction 139, 287–301 (2010). - PubMed
-
- Rathke C., Baarends W. M., Awe S. & Renkawitz-Pohl R. Chromatin dynamics during spermiogenesis. BBA–Gene Regul. Mech. 1839, 155–168 (2014). - PubMed
-
- Lewis J. D., Song Y., Jong M. E., Bagha S. M. & Ausió J. A walk though vertebrate and invertebrate protamines. Chromosoma 111, 473–482 (2003). - PubMed
-
- Gaucher J. et al.. From meiosis to postmeiotic events: the secrets of histone disappearance. FEBS J. 277, 599–604 (2010). - PubMed
-
- Eirin-López J. M. & Ausió J. Origin and evolution of chromosomal sperm proteins. Bioessays 31, 1062–1070 (2009). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
