

Time-Resolved Investigation of Molecular Components Involved in the Induction of [Formula: see text] High Affinity Transport System in Maize Roots
- PMID: 27877183
- PMCID: PMC5099785
- DOI: 10.3389/fpls.2016.01657
Time-Resolved Investigation of Molecular Components Involved in the Induction of [Formula: see text] High Affinity Transport System in Maize Roots
Abstract
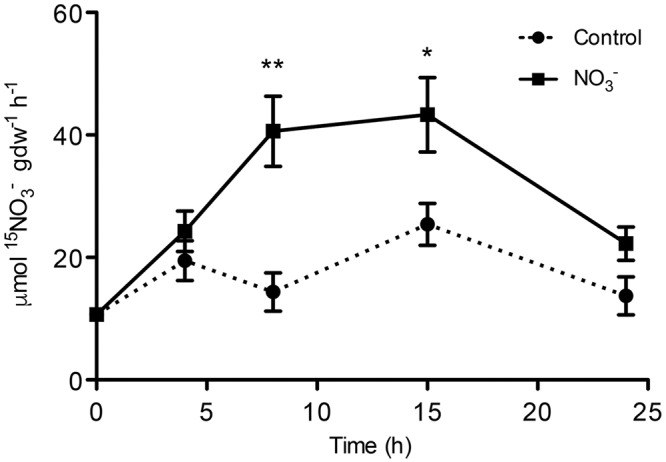
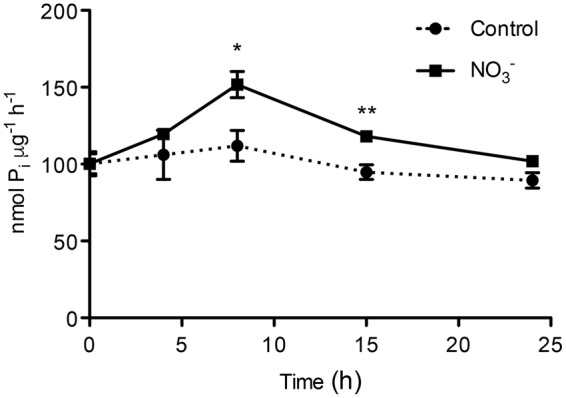
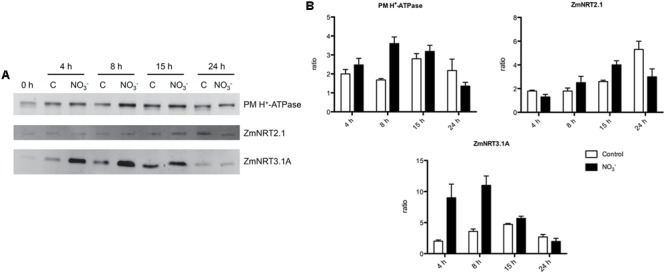
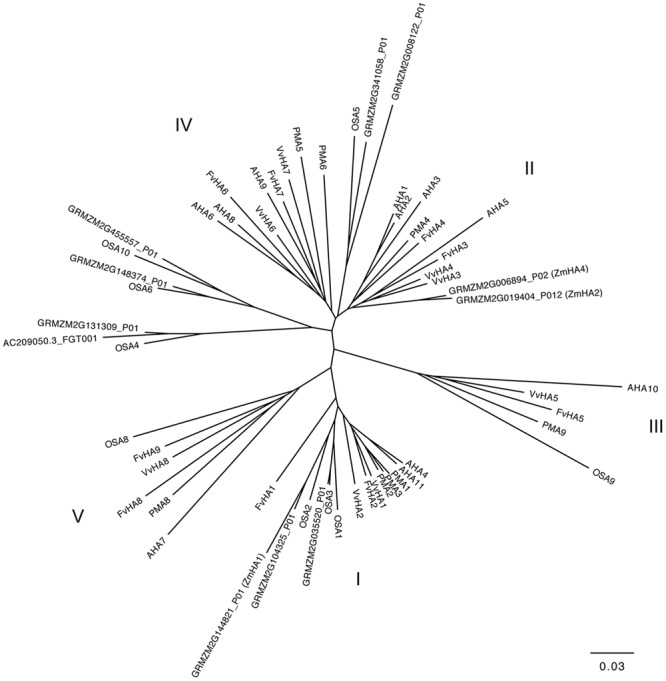
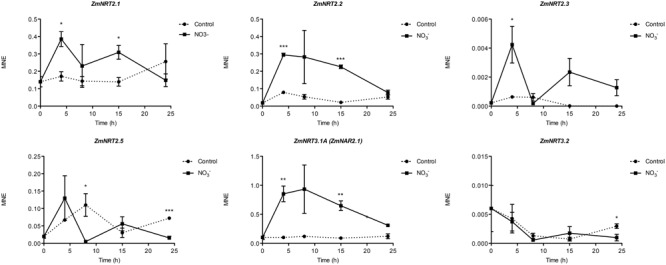
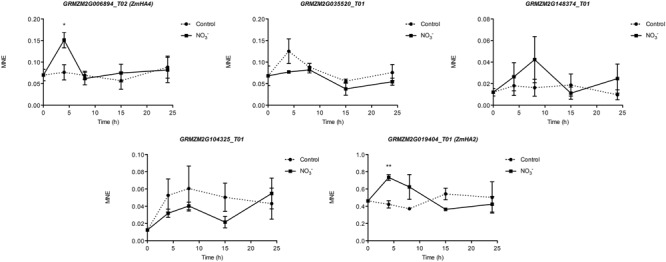
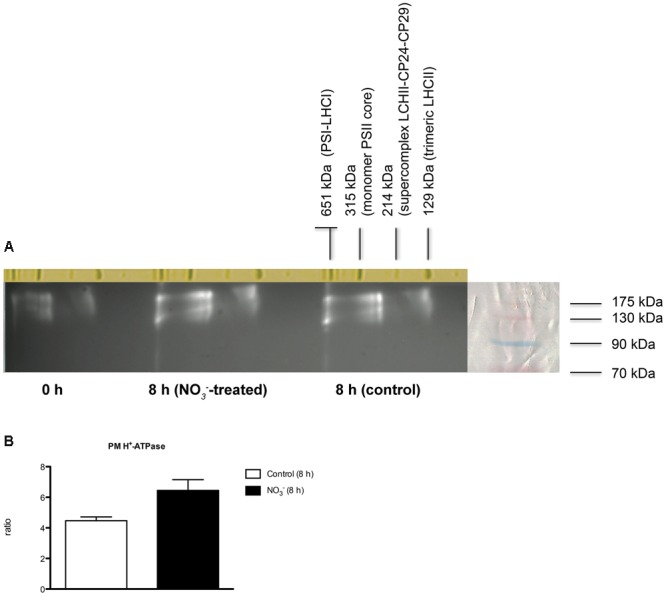
The induction, i.e., the rapid increase of nitrate ([Formula: see text]) uptake following the exposure of roots to the anion, was studied integrating physiological and molecular levels in maize roots. Responses to [Formula: see text] treatment were characterized in terms of changes in [Formula: see text] uptake rate and plasma membrane (PM) H+-ATPase activity and related to transcriptional and protein profiles of NRT2, NRT3, and PM H+-ATPase gene families. The behavior of transcripts and proteins of ZmNRT2s and ZmNRT3s suggested that the regulation of the activity of inducible high-affinity transport system (iHATS) is mainly based on the transcriptional/translational modulation of the accessory protein ZmNRT3.1A. Furthermore, ZmNRT2.1 and ZmNRT3.1A appear to be associated in a ∼150 kDa oligomer. The expression trend during the induction of the 11 identified PM H+-ATPase transcripts indicates that those mainly involved in the response to [Formula: see text] treatment are ZmHA2 and ZmHA4. Yet, partial correlation between the gene expression, protein levels and enzyme activity suggests an involvement of post-transcriptional and post-translational mechanisms of regulation. A non-denaturing Deriphat-PAGE approach allowed demonstrating for the first time that PM H+-ATPase can occur in vivo as hexameric complex together with the already described monomeric and dimeric forms.
Keywords: NO-3 induction; PM H+-ATPase; ZmNRT2.1; ZmNRT3.1A; maize; protein complexes.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources