

Downregulation of B-myb promotes senescence via the ROS-mediated p53/p21 pathway, in vascular endothelial cells
- PMID: 27878894
- PMCID: PMC6529094
- DOI: 10.1111/cpr.12319
Downregulation of B-myb promotes senescence via the ROS-mediated p53/p21 pathway, in vascular endothelial cells
Abstract
Objectives: To reveal whether B-myb is involved in preventing senescence of vascular endothelial cells, and if so, to identify possible mechanisms for it.
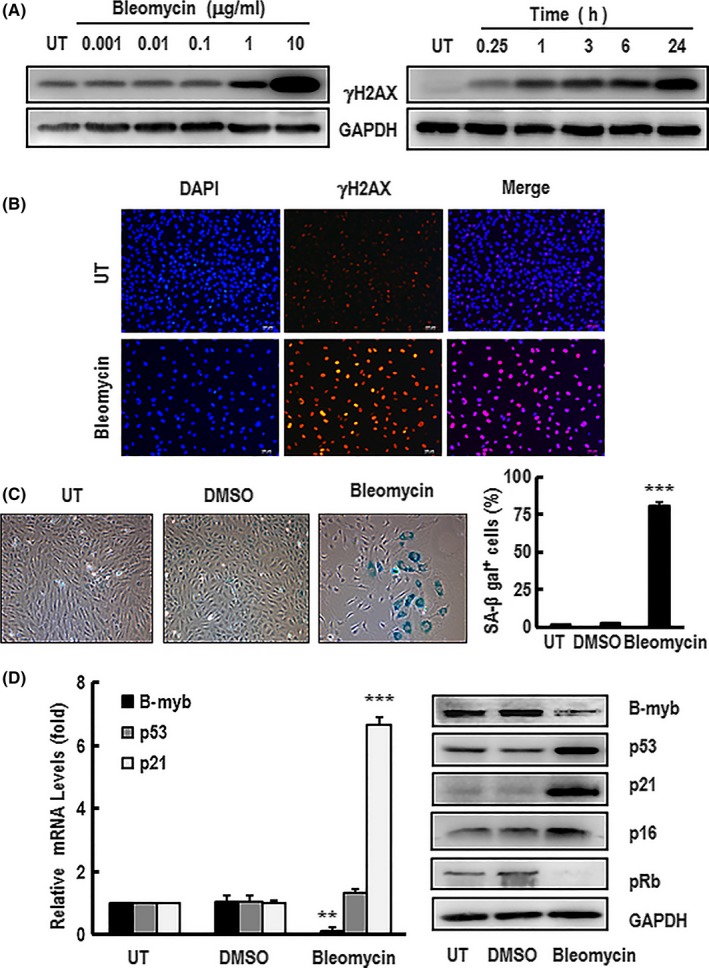
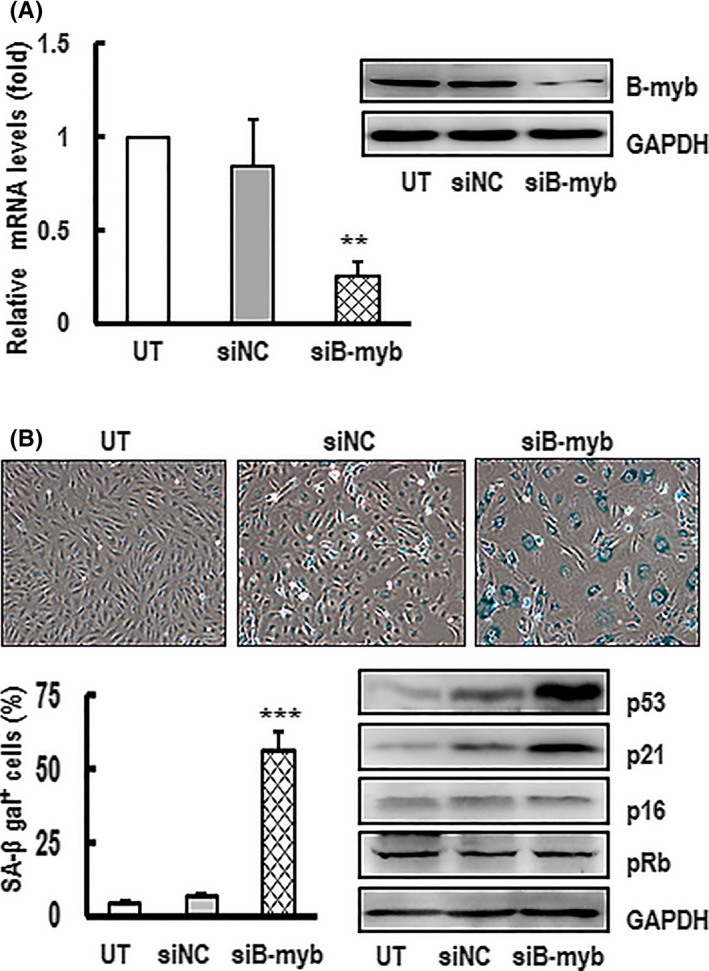
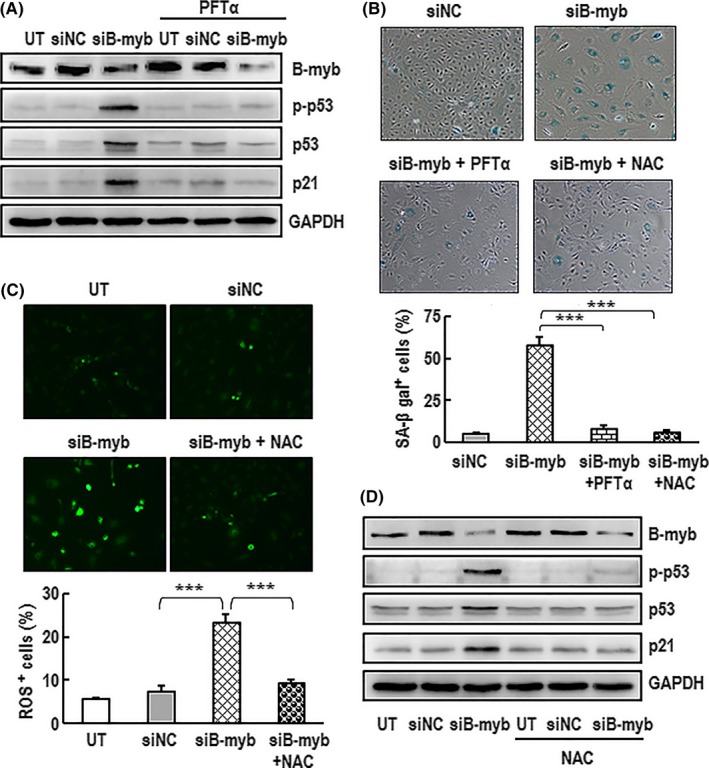
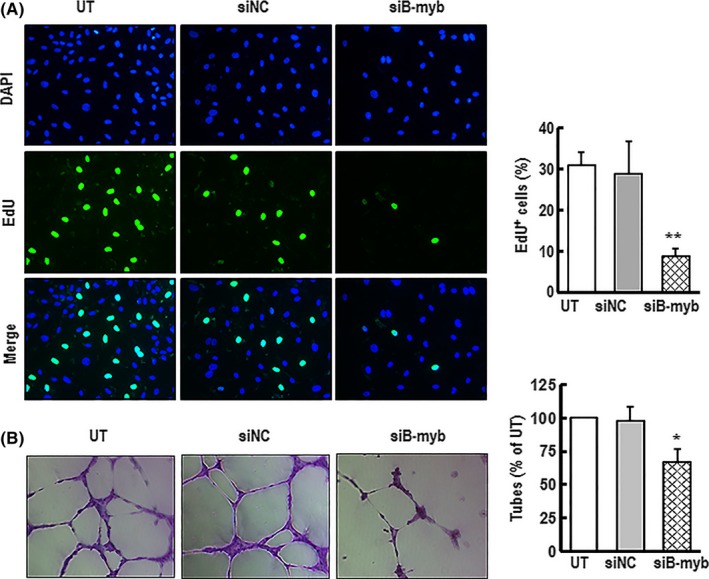
Materials and methods: C57/BL6 male mice and primary human aortic endothelial cells (HAECs) were used. Bleomycin was applied to induce stress-related premature senescence. B-myb knockdown was achieved using an siRNA technique and cell senescence was assessed using the senescence-associated β-galactosidase (SA-β-gal) assay. Intracellular reactive oxygen species (ROS) production was analysed using an ROS assay kit and cell proliferation was evaluated using KFluor488 EdU kit. Capillary tube network formation was determined by Matrigel assay. Expressions of mRNA and protein levels were detected by real-time PCR and western blotting.
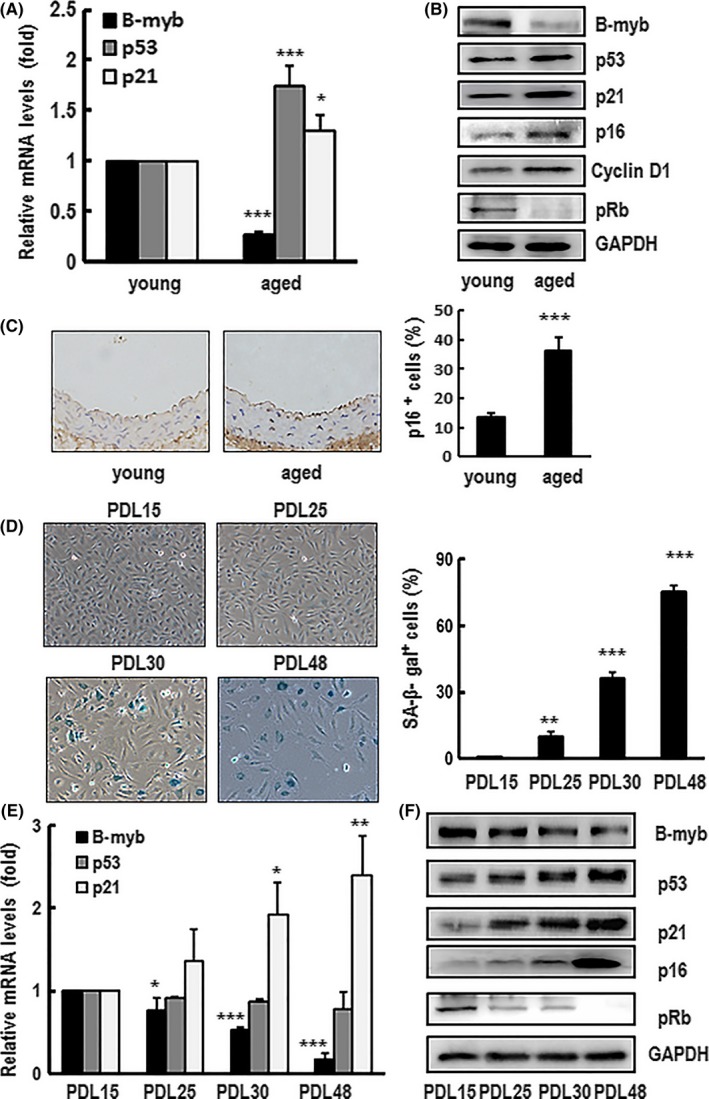
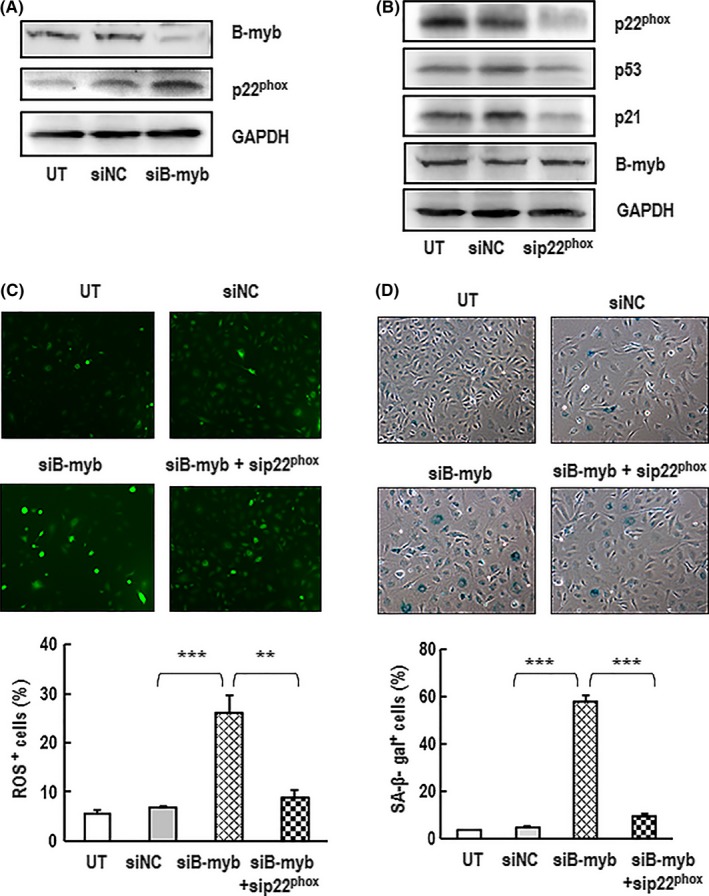
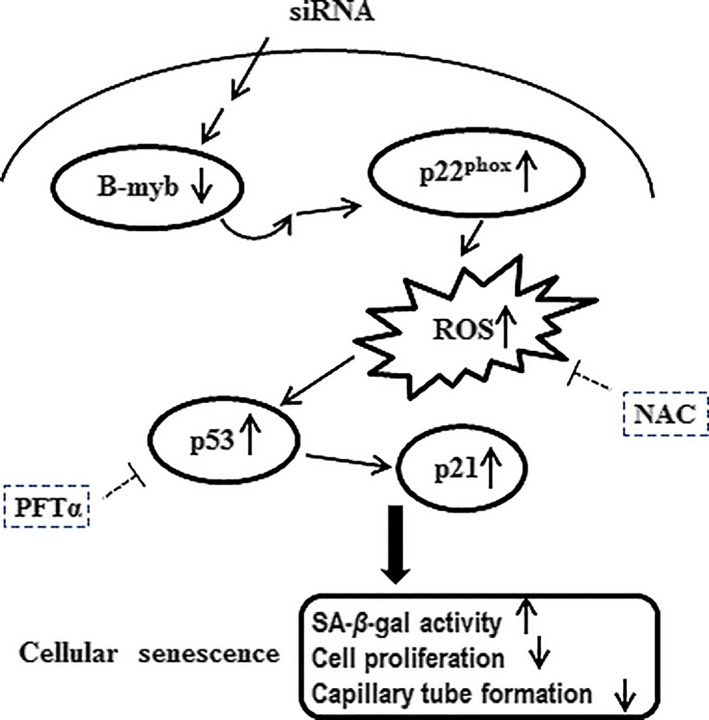
Results: B-myb expression significantly decreased, while p53 and p21 expressions increased in the aortas of aged mice. This expression pattern was also found in replicative senescent HAECs and senescent HAECs induced by bleomycin. B-myb knockdown resulted in upregulation of p22phox , ROS accumulation and cell senescence of HAECs. Downregulation of B-myb significantly inhibited cell proliferation and capillary tube network formation and activated the p53/p21 signalling pathway. Blocking ROS production or inhibiting p53 activation remarkably attenuated SA-β-gal activity and delayed cell senescence induced by B-myb-silencing.
Conclusion: Downregulation of B-myb induced senescence by upregulation of p22phox and activation of the ROS/p53/p21 pathway, in our vascular endothelial cells, suggesting that B-myb may be a novel candidate for regulating cell senescence to protect against endothelial senescence-related cardiovascular diseases.
© 2016 John Wiley & Sons Ltd.
Figures
Similar articles
-
GDF15 contributes to radiation-induced senescence through the ROS-mediated p16 pathway in human endothelial cells.Oncotarget. 2016 Mar 1;7(9):9634-44. doi: 10.18632/oncotarget.7457. Oncotarget. 2016. PMID: 26909594 Free PMC article.
-
UNC5B Promotes Vascular Endothelial Cell Senescence via the ROS-Mediated P53 Pathway.Oxid Med Cell Longev. 2021 Jun 20;2021:5546711. doi: 10.1155/2021/5546711. eCollection 2021. Oxid Med Cell Longev. 2021. PMID: 34239689 Free PMC article.
-
Interferon-gamma induces cellular senescence through p53-dependent DNA damage signaling in human endothelial cells.Mech Ageing Dev. 2009 Mar;130(3):179-88. doi: 10.1016/j.mad.2008.11.004. Epub 2008 Nov 21. Mech Ageing Dev. 2009. PMID: 19071156
-
Cellular senescence and aging: the role of B-MYB.Aging Cell. 2014 Oct;13(5):773-9. doi: 10.1111/acel.12242. Epub 2014 Jul 1. Aging Cell. 2014. PMID: 24981831 Free PMC article. Review.
-
B-Myb, cancer, senescence, and microRNAs.Cancer Res. 2011 Aug 15;71(16):5370-3. doi: 10.1158/0008-5472.CAN-11-1044. Epub 2011 Aug 9. Cancer Res. 2011. PMID: 21828240 Free PMC article. Review.
Cited by
-
Natural Polyphenols Targeting Senescence: A Novel Prevention and Therapy Strategy for Cancer.Int J Mol Sci. 2020 Jan 20;21(2):684. doi: 10.3390/ijms21020684. Int J Mol Sci. 2020. PMID: 31968672 Free PMC article. Review.
-
Histone Lysine Methylation Modification and Its Role in Vascular Calcification.Front Endocrinol (Lausanne). 2022 Jun 16;13:863708. doi: 10.3389/fendo.2022.863708. eCollection 2022. Front Endocrinol (Lausanne). 2022. PMID: 35784574 Free PMC article. Review.
-
ROS-mediated lysosomal membrane permeabilization and autophagy inhibition regulate bleomycin-induced cellular senescence.Autophagy. 2024 Sep;20(9):2000-2016. doi: 10.1080/15548627.2024.2353548. Epub 2024 May 18. Autophagy. 2024. PMID: 38762757 Free PMC article.
-
Fisetin ameliorates atherosclerosis by regulating PCSK9 and LOX-1 in apoE-/- mice.Exp Ther Med. 2021 Jan;21(1):25. doi: 10.3892/etm.2020.9457. Epub 2020 Nov 9. Exp Ther Med. 2021. PMID: 33262811 Free PMC article.
-
Syringin protects high glucose-induced BMSC injury, cell senescence, and osteoporosis by inhibiting JAK2/STAT3 signaling.J Appl Biomed. 2024 Dec;22(4):197-207. doi: 10.32725/jab.2024.021. Epub 2024 Oct 21. J Appl Biomed. 2024. PMID: 40033807
References
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
