

Brd4 Activates Early Viral Transcription upon Human Papillomavirus 18 Infection of Primary Keratinocytes
- PMID: 27879331
- PMCID: PMC5120138
- DOI: 10.1128/mBio.01644-16
Brd4 Activates Early Viral Transcription upon Human Papillomavirus 18 Infection of Primary Keratinocytes
Abstract
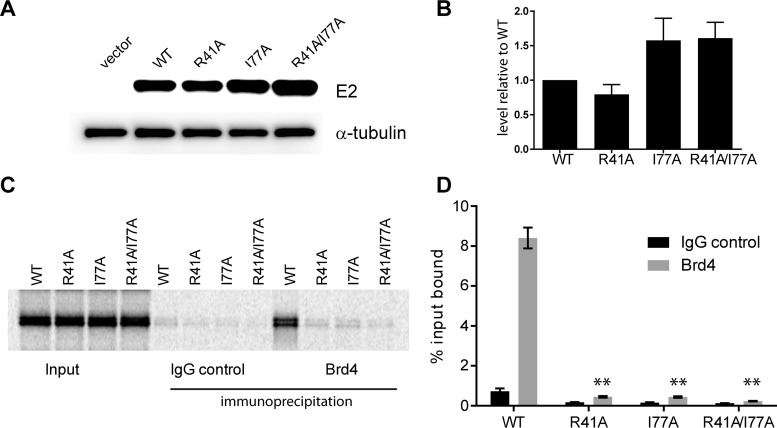
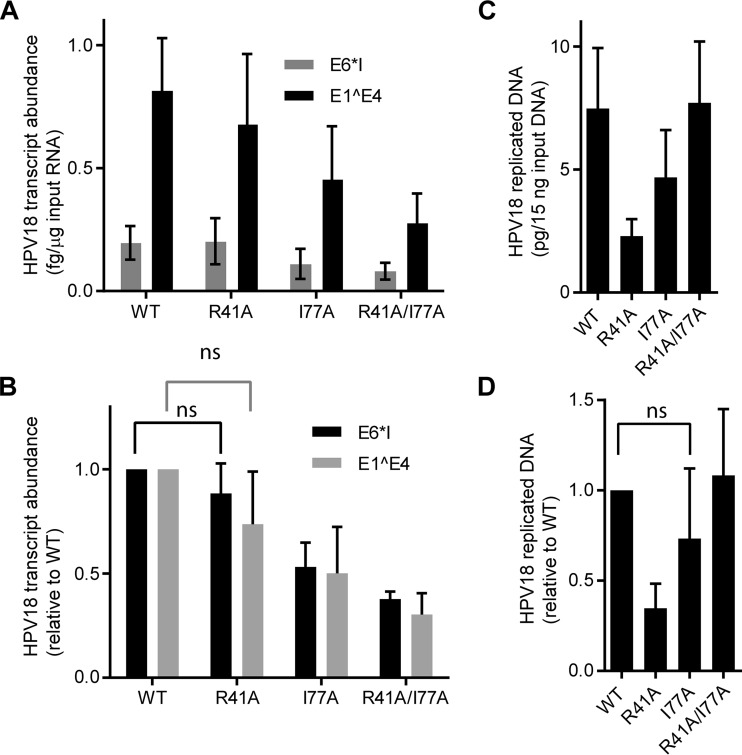
Human papillomaviruses (HPVs) replicate in the cutaneous and mucosal epithelia, and the infectious cycle is synchronous with the differentiation program of the host keratinocytes. The virus initially infects dividing cells in the lower layers of the epithelium, where it establishes a persistent infection. The viral genome is maintained as a low-copy-number, extrachromosomal element in these proliferating cells but switches to the late stage of the life cycle in differentiated cells. The cellular chromatin adaptor protein Brd4 is involved in several stages and processes of the viral life cycle. In concert with the viral transcriptional regulator E2, Brd4 can repress transcription from the early viral promoter. Brd4 and E2 form a complex with the viral genome that associates with host chromosomes to partition the viral genome in dividing cells; Brd4 also localizes to active sites of productive HPV DNA replication. However, because of the difficulties in producing HPV viral particles, the role of Brd4 in modulating viral transcription and replication at the initial stage of infection is unclear. In this study, we have used an HPV18 quasivirus-based genome delivery system to assess the role of Brd4 in the initial infectivity of primary human keratinocytes. We show that, upon infection of primary human keratinocytes with HPV18 quasivirus, Brd4 activates viral transcription and replication. Furthermore, this activation is independent of the functional interaction between Brd4 and the HPV18 E2 protein.
Importance: HPVs lack encapsidated proteins and so rely exquisitely on host cellular factors to initiate their gene expression programs in newly infected cells. Brd4 is an important cellular chromatin adaptor molecule that normally activates host transcription initiation and elongation. In this study, we further optimize and utilize a quasivirus infection system to show that Brd4 activates HPV18 transcription at early stages of infection. HPVs are important human pathogens causing a wide range of cutaneous and tumorigenic morbidities. Therefore, specifically targeting this protein could provide a new target of therapeutic prevention of establishment of HPV infections.
Copyright © 2016 McKinney et al.
Figures
References
-
- Yang EJ, Quick MC, Hanamornroongruang S, Lai K, Doyle LA, McKeon FD, Xian W, Crum CP, Herfs M. 2015. Microanatomy of the cervical and anorectal squamocolumnar junctions: a proposed model for anatomical differences in HPV-related cancer risk. Mod Pathol 28:994–1000. doi: 10.1038/modpathol.2015.54. - DOI - PMC - PubMed
-
- Aydin I, Weber S, Snijder B, Samperio Ventayol P, Kühbacher A, Becker M, Day PM, Schiller JT, Kann M, Pelkmans L, Helenius A, Schelhaas M. 2014. Large scale RNAi reveals the requirement of nuclear envelope breakdown for nuclear import of human papillomaviruses. PLoS Pathog 10:e1004162. doi: 10.1371/journal.ppat.1004162. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
