

Characterization of Class III Peroxidases from Switchgrass
- PMID: 27879392
- PMCID: PMC5210742
- DOI: 10.1104/pp.16.01426
Characterization of Class III Peroxidases from Switchgrass
Abstract
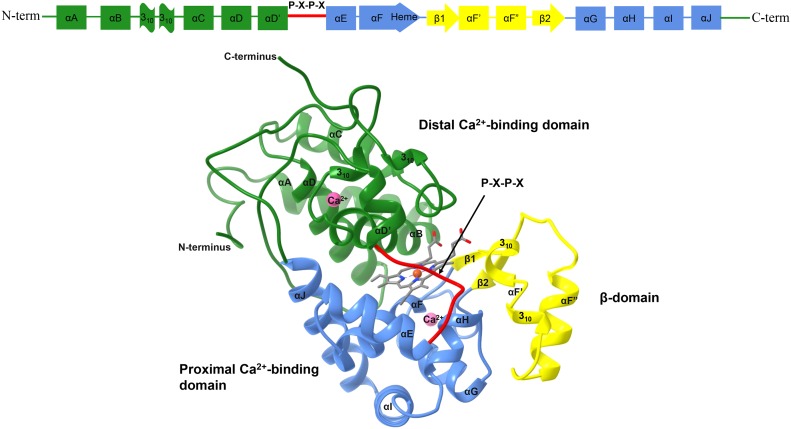
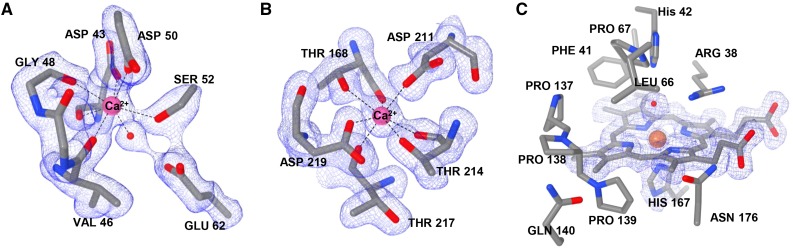
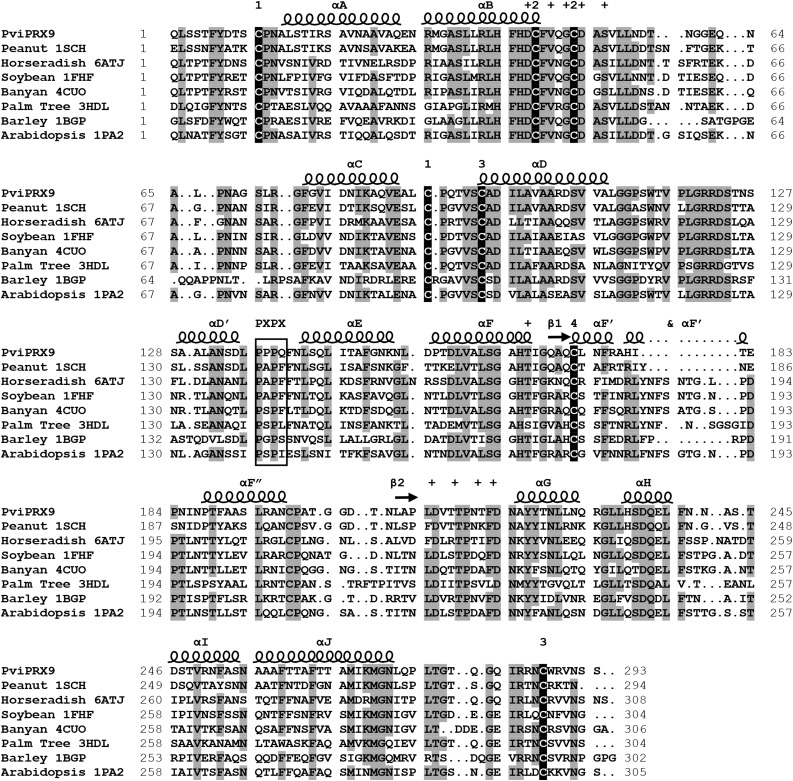
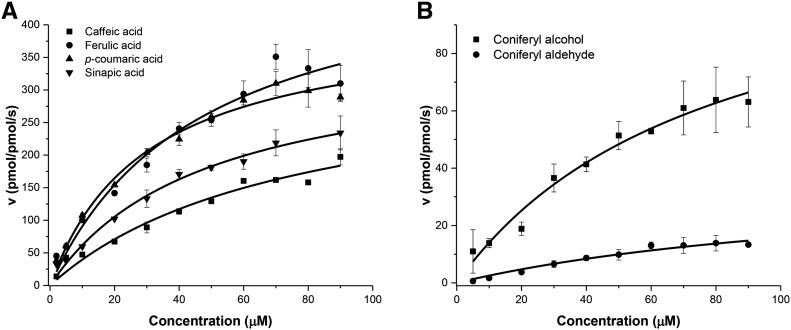
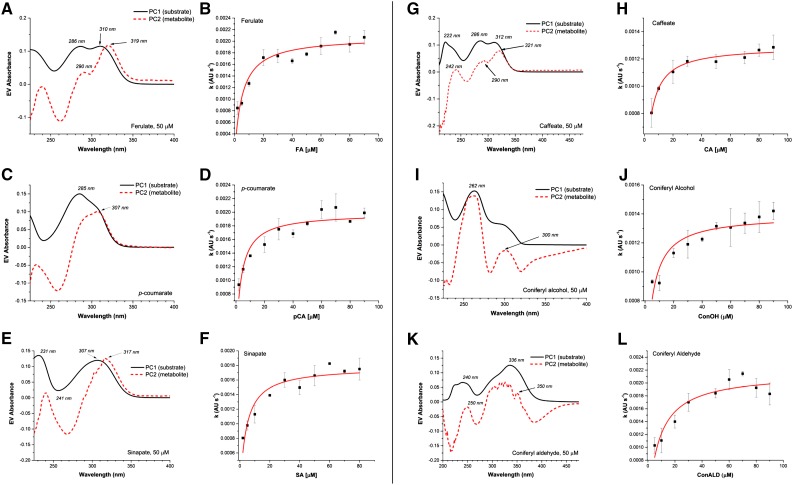
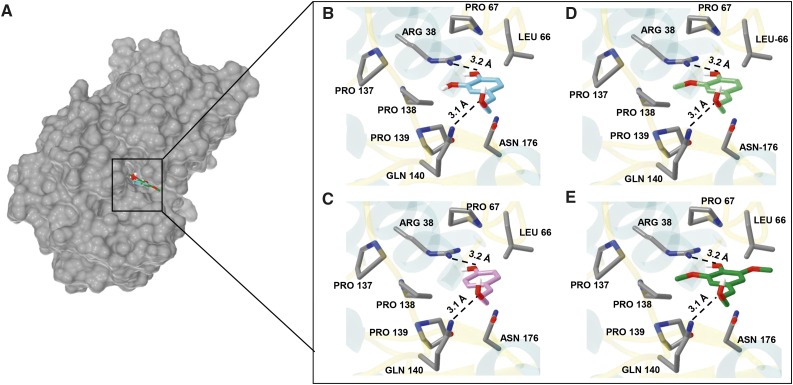
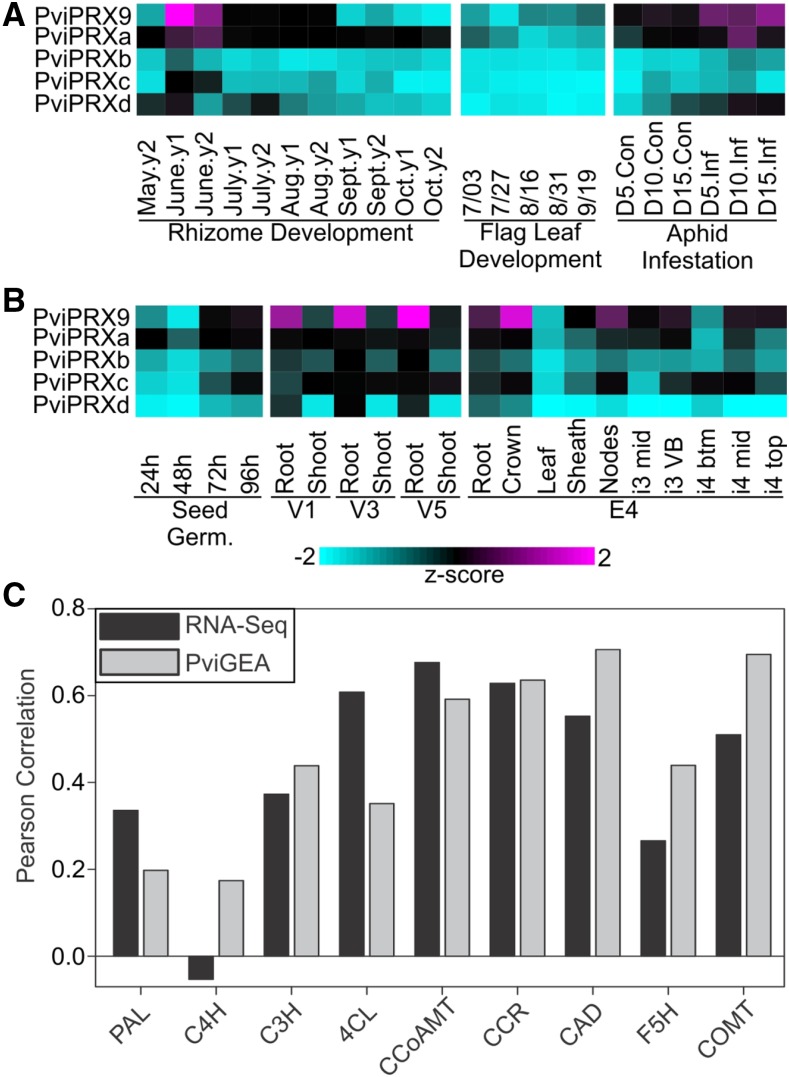
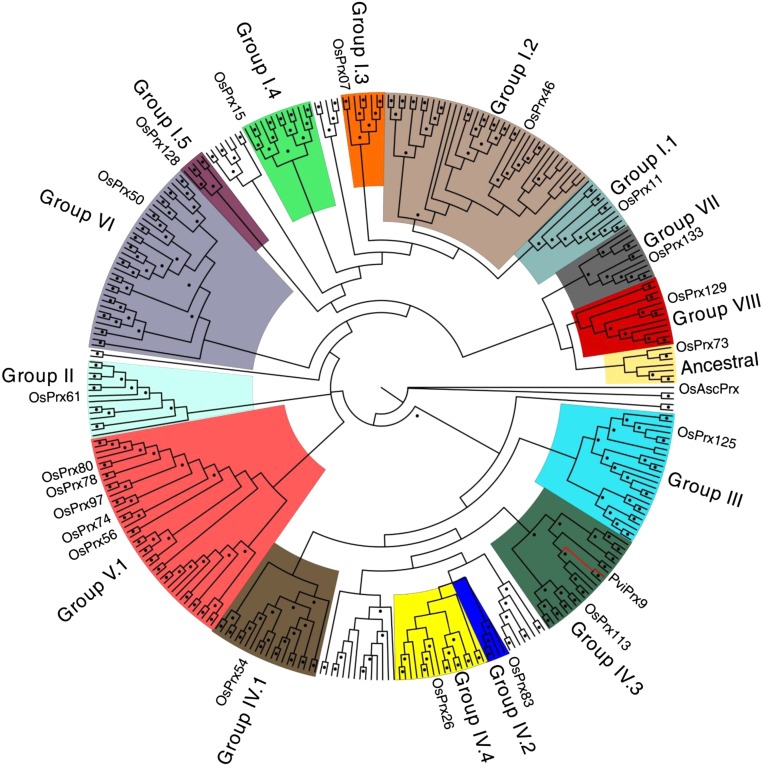
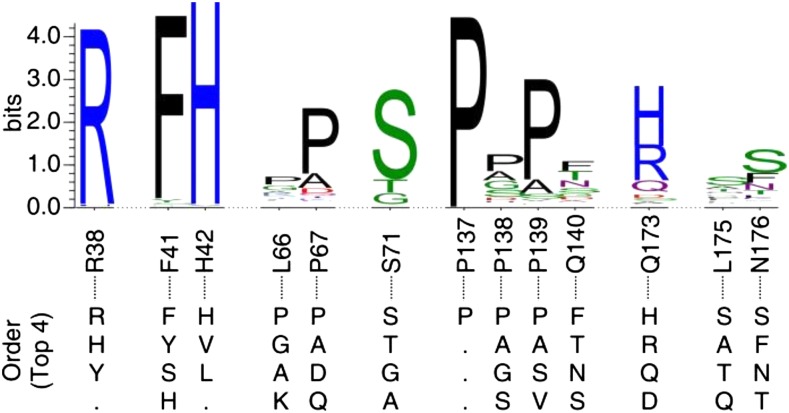
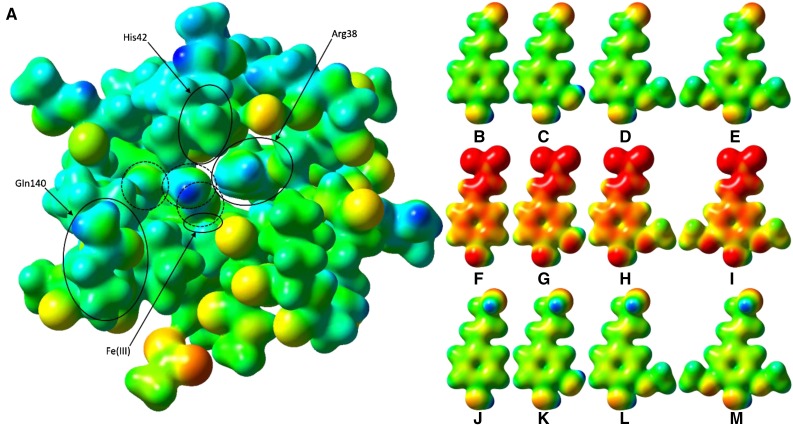
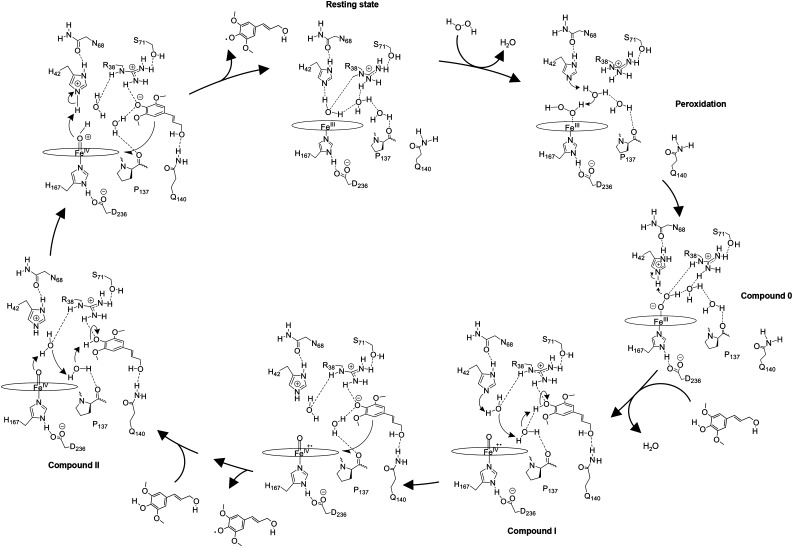
Class III peroxidases (CIIIPRX) catalyze the oxidation of monolignols, generate radicals, and ultimately lead to the formation of lignin. In general, CIIIPRX genes encode a large number of isozymes with ranges of in vitro substrate specificities. In order to elucidate the mode of substrate specificity of these enzymes, we characterized one of the CIIIPRXs (PviPRX9) from switchgrass (Panicum virgatum), a strategic plant for second-generation biofuels. The crystal structure, kinetic experiments, molecular docking, as well as expression patterns of PviPRX9 across multiple tissues and treatments, along with its levels of coexpression with the majority of genes in the monolignol biosynthesis pathway, revealed the function of PviPRX9 in lignification. Significantly, our study suggested that PviPRX9 has the ability to oxidize a broad range of phenylpropanoids with rather similar efficiencies, which reflects its role in the fortification of cell walls during normal growth and root development and in response to insect feeding. Based on the observed interactions of phenylpropanoids in the active site and analysis of kinetics, a catalytic mechanism involving two water molecules and residues histidine-42, arginine-38, and serine-71 was proposed. In addition, proline-138 and gluntamine-140 at the 137P-X-P-X140 motif, leucine-66, proline-67, and asparagine-176 may account for the broad substrate specificity of PviPRX9. Taken together, these observations shed new light on the function and catalysis of PviPRX9 and potentially benefit efforts to improve biomass conservation properties in bioenergy and forage crops.
© 2017 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Abelskov AK, Smith AT, Rasmussen CB, Dunford HB, Welinder KG (1997) pH dependence and structural interpretation of the reactions of Coprinus cinereus peroxidase with hydrogen peroxide, ferulic acid, and 2,2′-azinobis(3-ethylbenzthiazoline-6-sulfonic acid). Biochemistry 36: 9453–9463 - PubMed
-
- Almagro L, Gómez Ros LV, Belchi-Navarro S, Bru R, Ros Barceló A, Pedreño MA (2009) Class III peroxidases in plant defence reactions. J Exp Bot 60: 377–390 - PubMed
-
- Baek HK, Van Wart HE (1989) Elementary steps in the formation of horseradish peroxidase compound I: direct observation of compound 0, a new intermediate with a hyperporphyrin spectrum. Biochemistry 28: 5714–5719 - PubMed
-
- Bakovic M, Dunford HB (1993) Kinetics of the oxidation of p-coumaric acid by prostaglandin H synthase and hydrogen peroxide. Biochemistry 32: 833–840 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
