

Hyperglycemia induces defective Ca2+ homeostasis in cardiomyocytes
- PMID: 27881388
- PMCID: PMC5283918
- DOI: 10.1152/ajpheart.00737.2016
Hyperglycemia induces defective Ca2+ homeostasis in cardiomyocytes
Abstract
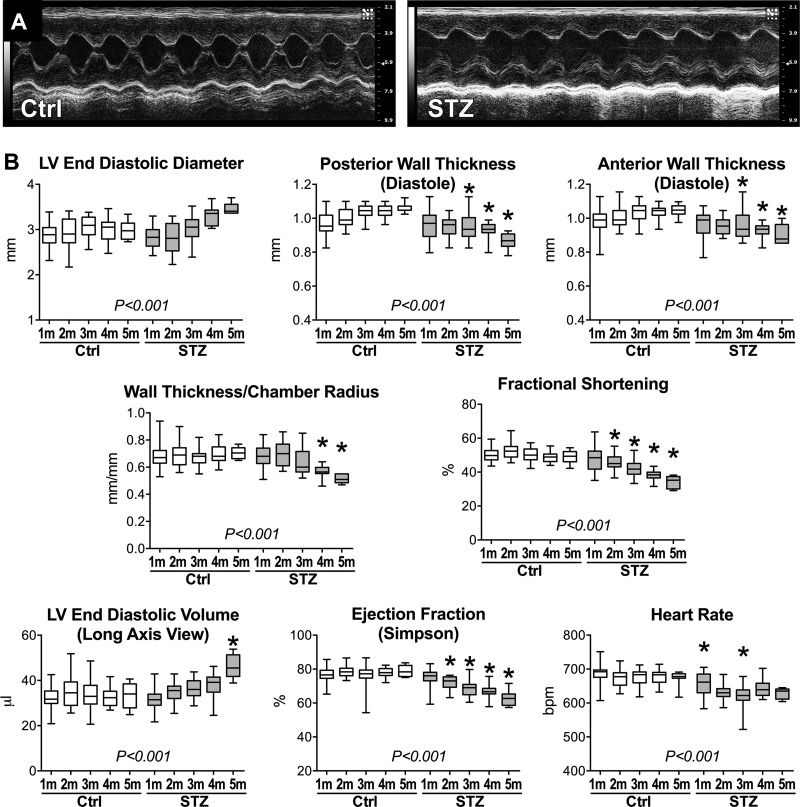
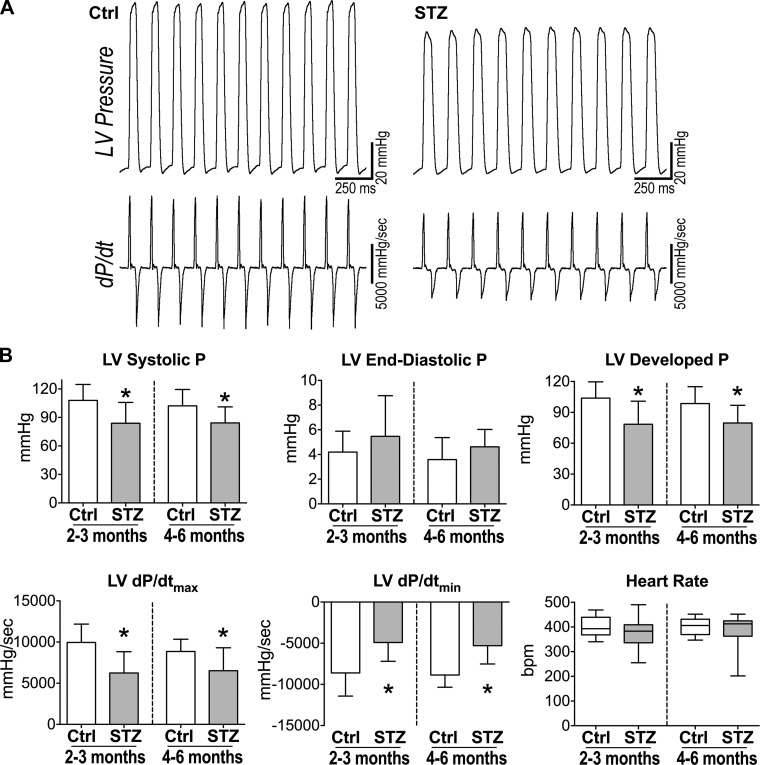
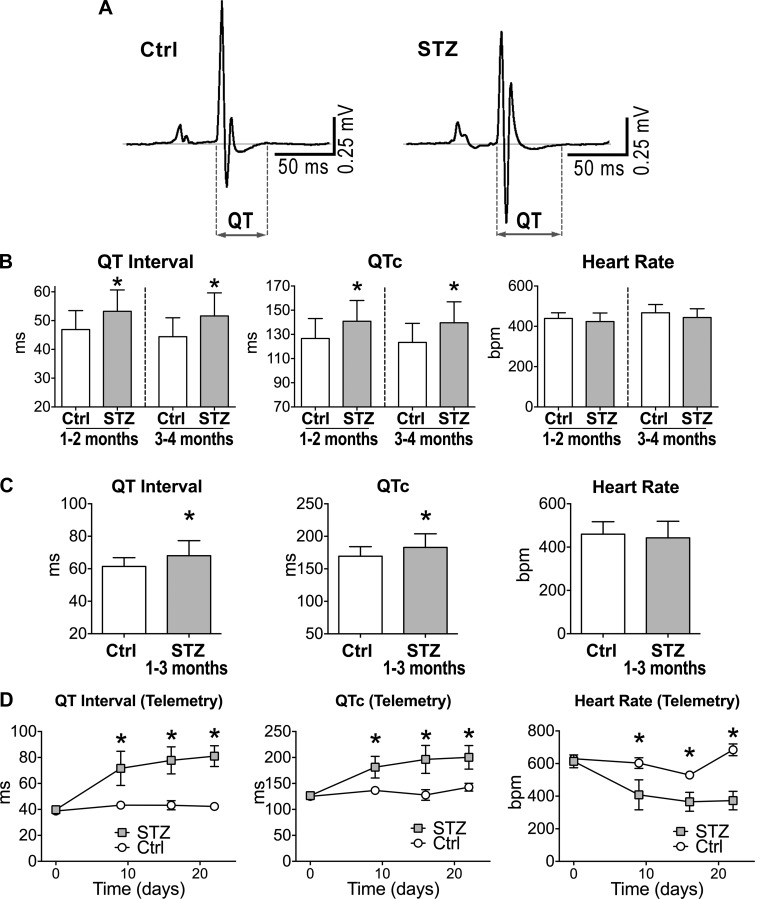
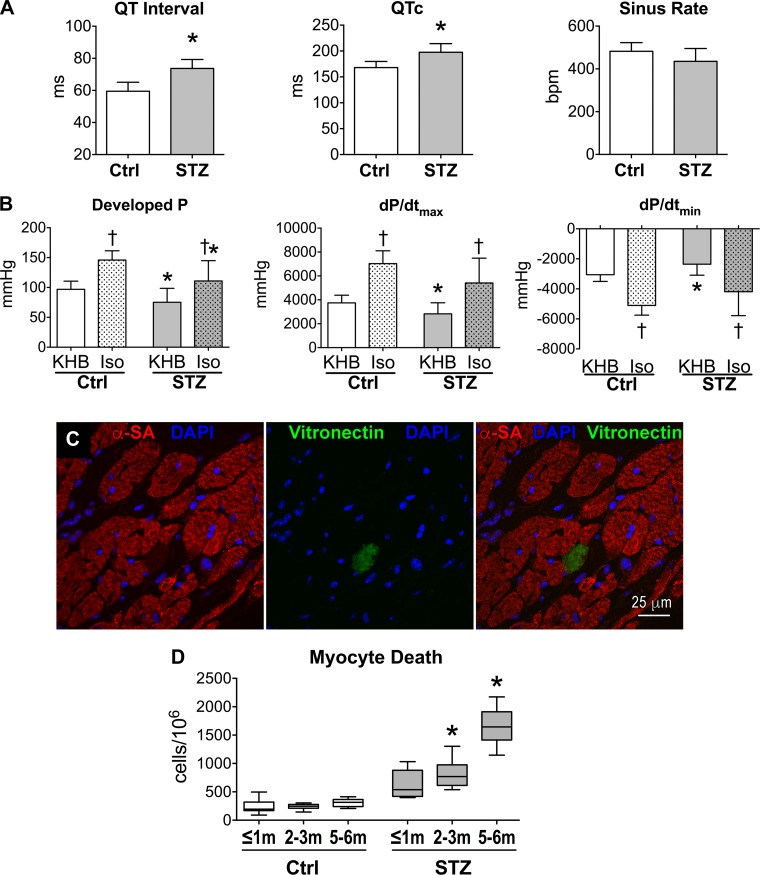
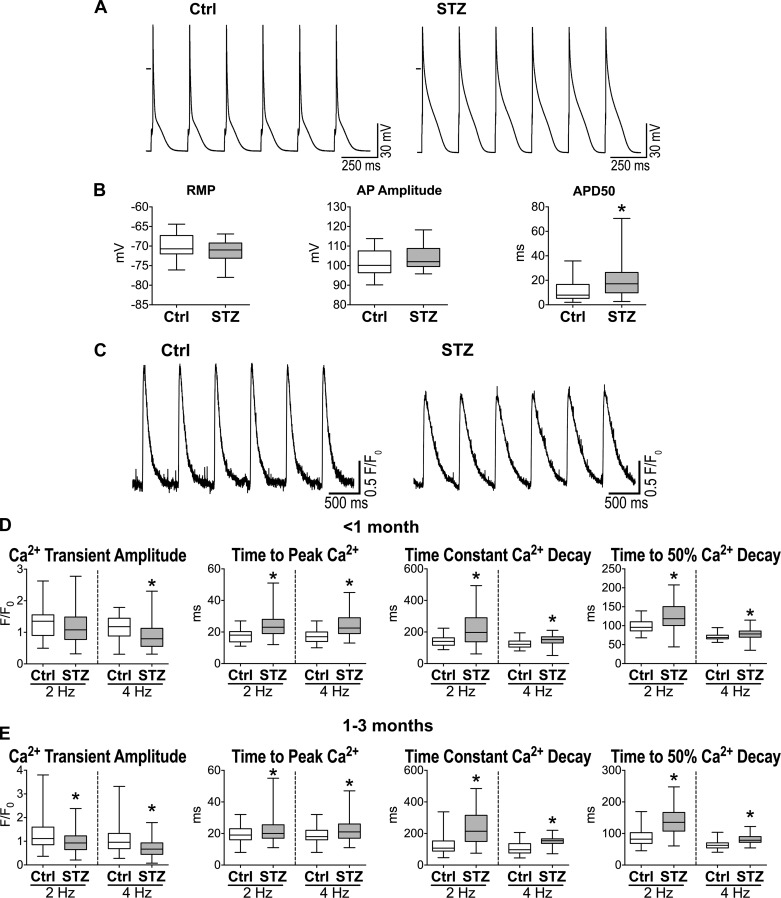
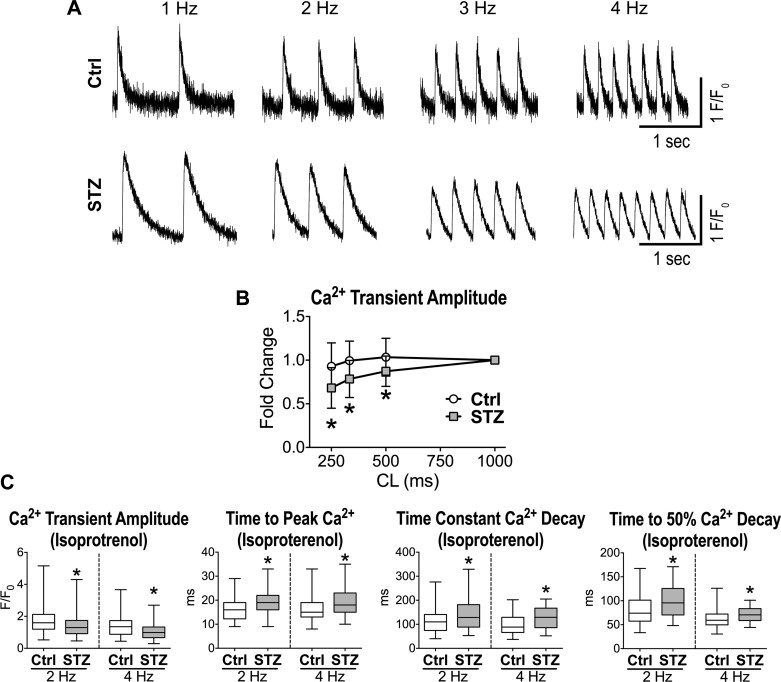
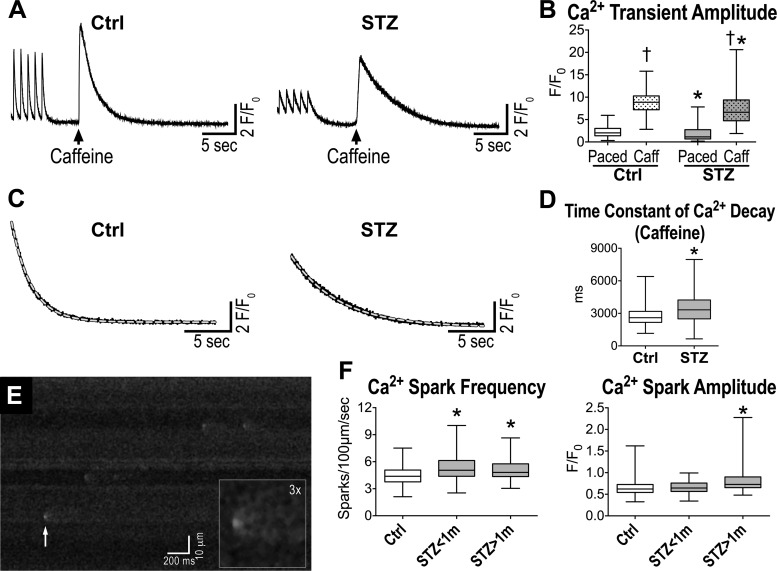
Diabetes and other metabolic conditions characterized by elevated blood glucose constitute important risk factors for cardiovascular disease. Hyperglycemia targets myocardial cells rendering ineffective mechanical properties of the heart, but cellular alterations dictating the progressive deterioration of cardiac function with metabolic disorders remain to be clarified. In the current study, we examined the effects of hyperglycemia on cardiac function and myocyte physiology by employing mice with high blood glucose induced by administration of streptozotocin, a compound toxic to insulin-producing β-cells. We found that hyperglycemia initially delayed the electrical recovery of the heart, whereas cardiac function became defective only after ~2 mo with this condition and gradually worsened with time. Prolonged hyperglycemia was associated with increased chamber dilation, thinning of the left ventricle (LV), and myocyte loss. Cardiomyocytes from hyperglycemic mice exhibited defective Ca2+ transients before the appearance of LV systolic defects. Alterations in Ca2+ transients involved enhanced spontaneous Ca2+ releases from the sarcoplasmic reticulum (SR), reduced cytoplasmic Ca2+ clearance, and declined SR Ca2+ load. These defects have important consequences on myocyte contraction, relaxation, and mechanisms of rate adaptation. Collectively, our data indicate that hyperglycemia alters intracellular Ca2+ homeostasis in cardiomyocytes, hindering contractile activity and contributing to the manifestation of the diabetic cardiomyopathy.
New & noteworthy: We have investigated the effects of hyperglycemia on cardiomyocyte physiology and ventricular function. Our results indicate that defective Ca2+ handling is a critical component of the progressive deterioration of cardiac performance of the diabetic heart.
Keywords: Ca2+ handling; diabetes; myocytes; ventricular function.
Copyright © 2017 the American Physiological Society.
Figures
Similar articles
-
ß-blocker timolol prevents arrhythmogenic Ca²⁺ release and normalizes Ca²⁺ and Zn²⁺ dyshomeostasis in hyperglycemic rat heart.PLoS One. 2013 Jul 29;8(7):e71014. doi: 10.1371/journal.pone.0071014. Print 2013. PLoS One. 2013. PMID: 23923043 Free PMC article.
-
Regional effects of streptozotocin-induced diabetes on shortening and calcium transport in epicardial and endocardial myocytes from rat left ventricle.Physiol Rep. 2016 Nov;4(22):e13034. doi: 10.14814/phy2.13034. Physiol Rep. 2016. PMID: 27884956 Free PMC article.
-
Interval training normalizes cardiomyocyte function, diastolic Ca2+ control, and SR Ca2+ release synchronicity in a mouse model of diabetic cardiomyopathy.Circ Res. 2009 Sep 11;105(6):527-36. doi: 10.1161/CIRCRESAHA.109.199810. Epub 2009 Aug 13. Circ Res. 2009. PMID: 19679837
-
Intracellular calcium levels are unchanged in the diabetic heart.Cardiovasc Res. 1997 Apr;34(1):91-8. doi: 10.1016/s0008-6363(97)00034-5. Cardiovasc Res. 1997. PMID: 9217877 Review.
-
Mechanisms of subcellular remodeling in heart failure due to diabetes.Heart Fail Rev. 2014 Jan;19(1):87-99. doi: 10.1007/s10741-013-9385-8. Heart Fail Rev. 2014. PMID: 23436108 Review.
Cited by
-
Ca2+ mishandling and mitochondrial dysfunction: a converging road to prediabetic and diabetic cardiomyopathy.Pflugers Arch. 2022 Jan;474(1):33-61. doi: 10.1007/s00424-021-02650-y. Epub 2022 Jan 3. Pflugers Arch. 2022. PMID: 34978597 Free PMC article. Review.
-
Diabetic Cardiomyopathy: Current and Future Therapies. Beyond Glycemic Control.Front Physiol. 2018 Oct 30;9:1514. doi: 10.3389/fphys.2018.01514. eCollection 2018. Front Physiol. 2018. PMID: 30425649 Free PMC article. Review.
-
Heart Rate Variability Reveals Altered Autonomic Regulation in Response to Myocardial Infarction in Experimental Animals.Front Cardiovasc Med. 2022 May 2;9:843144. doi: 10.3389/fcvm.2022.843144. eCollection 2022. Front Cardiovasc Med. 2022. PMID: 35586660 Free PMC article.
-
The Histone Deacetylase Inhibitor Suberoylanilide Hydroxamic Acid (SAHA) Restores Cardiomyocyte Contractility in a Rat Model of Early Diabetes.Int J Mol Sci. 2019 Apr 16;20(8):1873. doi: 10.3390/ijms20081873. Int J Mol Sci. 2019. PMID: 31014028 Free PMC article.
-
The cAMP/PKA signaling pathway conditions cardiac performance in experimental animals with metabolic syndrome.J Mol Cell Cardiol. 2024 Nov;196:35-51. doi: 10.1016/j.yjmcc.2024.09.002. Epub 2024 Sep 7. J Mol Cell Cardiol. 2024. PMID: 39251059
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous