

Environmental Pseudomonads Inhibit Cystic Fibrosis Patient-Derived Pseudomonas aeruginosa
- PMID: 27881418
- PMCID: PMC5203635
- DOI: 10.1128/AEM.02701-16
Environmental Pseudomonads Inhibit Cystic Fibrosis Patient-Derived Pseudomonas aeruginosa
Abstract
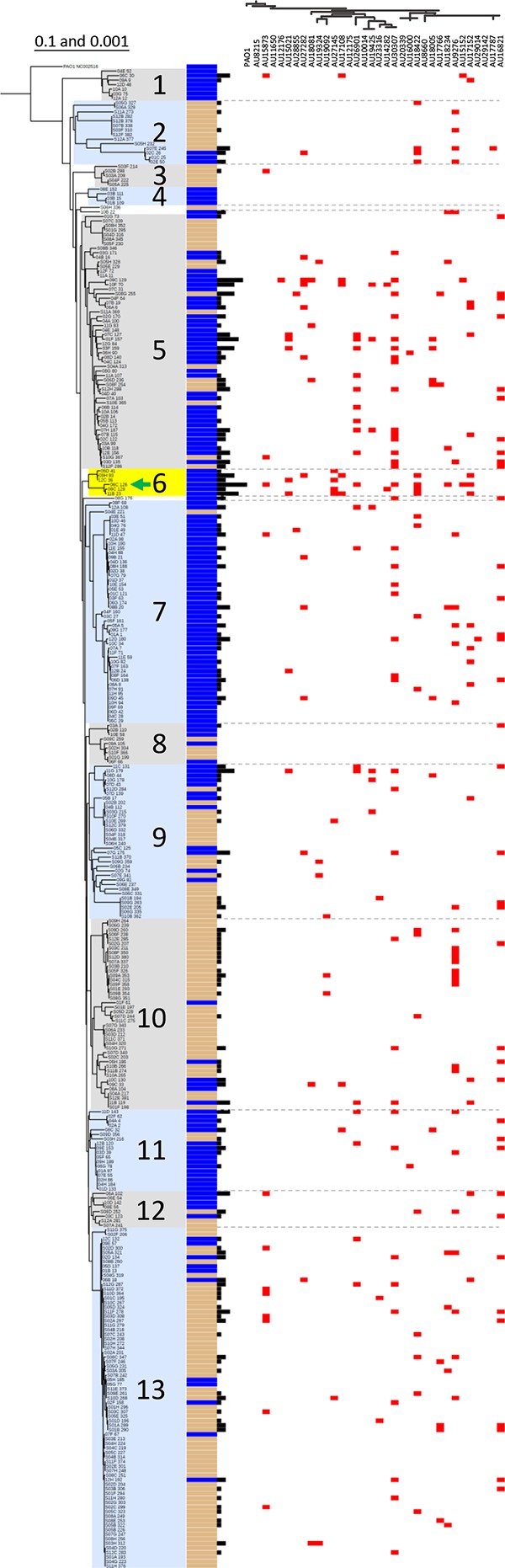
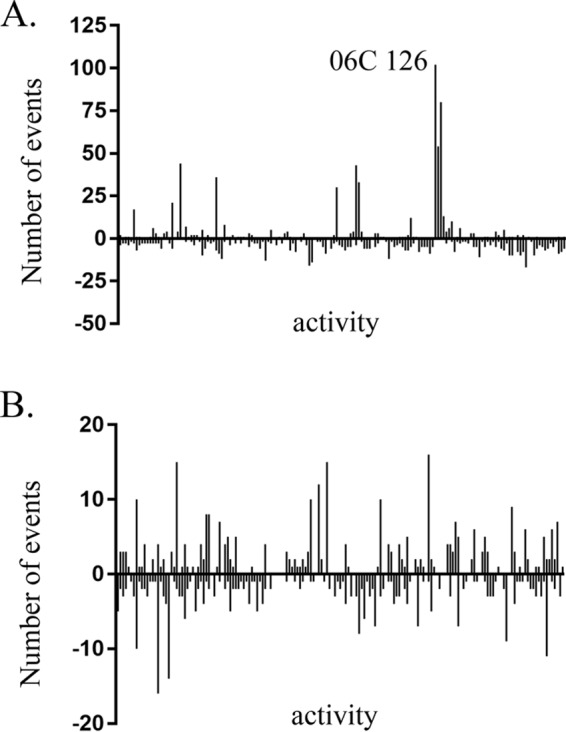
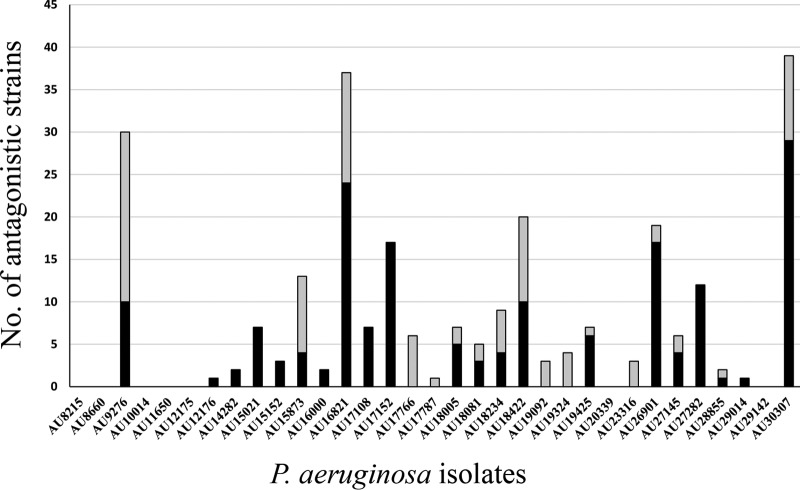
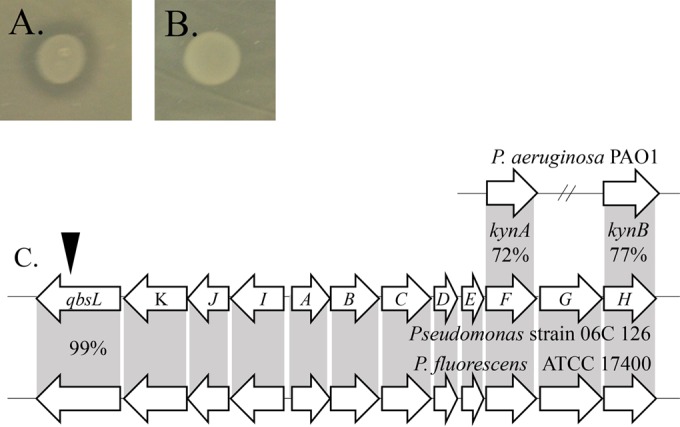
Pseudomonas aeruginosa is an opportunistic pathogen which is evolving resistance to many currently used antibiotics. While much research has been devoted to the roles of pathogenic P. aeruginosa in cystic fibrosis (CF) patients, less is known of its ecological properties. P. aeruginosa dominates the lungs during chronic infection in CF patients, yet its abundance in some environments is less than that of other diverse groups of pseudomonads. Here, we sought to determine if clinical isolates of P. aeruginosa are vulnerable to environmental pseudomonads that dominate soil and water habitats in one-to-one competitions which may provide a source of inhibitory factors. We isolated a total of 330 pseudomonads from diverse habitats of soil and freshwater ecosystems and competed these strains against one another to determine their capacity for antagonistic activity. Over 900 individual inhibitory events were observed. Extending the analysis to P. aeruginosa isolates revealed that clinical isolates, including ones with increased alginate production, were susceptible to competition by multiple environmental strains. We performed transposon mutagenesis on one isolate and identified an ∼14.8-kb locus involved in antagonistic activity. Only two other environmental isolates were observed to carry the locus, suggesting the presence of additional unique compounds or interactions among other isolates involved in outcompeting P. aeruginosa This collection of strains represents a source of compounds that are active against multiple pathogenic strains. With the evolution of resistance of P. aeruginosa to currently used antibiotics, these environmental strains provide opportunities for novel compound discovery against drug-resistant clinical strains.
Importance: We demonstrate that clinical CF-derived isolates of P. aeruginosa are susceptible to competition in the presence of environmental pseudomonads. We observed that many diverse environmental strains exhibited varied antagonistic profiles against a panel of clinical P. aeruginosa isolates, suggesting the presence of distinct mechanisms of inhibition among these ecological strains. Understanding the properties of these antagonistic events offers the potential for discoveries of antimicrobial compounds or metabolic pathways important to the development of novel treatments for P. aeruginosa infections.
Keywords: Pseudomonas aeruginosa; antagonistic; kynurenine; pseudomonads; siderophore; thioquinolobactin.
Copyright © 2016 Chatterjee et al.
Figures

Similar articles
-
Loci Encoding Compounds Potentially Active against Drug-Resistant Pathogens amidst a Decreasing Pool of Novel Antibiotics.Appl Environ Microbiol. 2019 Nov 14;85(23):e01438-19. doi: 10.1128/AEM.01438-19. Print 2019 Dec 1. Appl Environ Microbiol. 2019. PMID: 31540982 Free PMC article.
-
Competition in Biofilms between Cystic Fibrosis Isolates of Pseudomonas aeruginosa Is Shaped by R-Pyocins.mBio. 2019 Jan 29;10(1):e01828-18. doi: 10.1128/mBio.01828-18. mBio. 2019. PMID: 30696740 Free PMC article.
-
Genotypic and Phenotypic Diversity of Staphylococcus aureus Isolates from Cystic Fibrosis Patient Lung Infections and Their Interactions with Pseudomonas aeruginosa.mBio. 2020 Jun 23;11(3):e00735-20. doi: 10.1128/mBio.00735-20. mBio. 2020. PMID: 32576671 Free PMC article.
-
Pseudomonas aeruginosa chromosomal beta-lactamase in patients with cystic fibrosis and chronic lung infection. Mechanism of antibiotic resistance and target of the humoral immune response.APMIS Suppl. 2003;(116):1-47. APMIS Suppl. 2003. PMID: 14692154 Review.
-
Microevolution of Pseudomonas aeruginosa to a chronic pathogen of the cystic fibrosis lung.Curr Top Microbiol Immunol. 2013;358:91-118. doi: 10.1007/82_2011_199. Curr Top Microbiol Immunol. 2013. PMID: 22311171 Review.
Cited by
-
High Occurrence of Bacterial Competition Among Clinically Documented Opportunistic Pathogens Including Achromobacter xylosoxidans in Cystic Fibrosis.Front Microbiol. 2020 Sep 10;11:558160. doi: 10.3389/fmicb.2020.558160. eCollection 2020. Front Microbiol. 2020. PMID: 33013789 Free PMC article.
-
Multichannel Microfluidic Platform for Temporal-Spatial Investigation of Niche Roles of Pseudomonas aeruginosa and Escherichia coli within a Dual-Species Biofilm.Appl Environ Microbiol. 2023 Jul 26;89(7):e0065123. doi: 10.1128/aem.00651-23. Epub 2023 Jun 29. Appl Environ Microbiol. 2023. PMID: 37382537 Free PMC article.
-
Loci Encoding Compounds Potentially Active against Drug-Resistant Pathogens amidst a Decreasing Pool of Novel Antibiotics.Appl Environ Microbiol. 2019 Nov 14;85(23):e01438-19. doi: 10.1128/AEM.01438-19. Print 2019 Dec 1. Appl Environ Microbiol. 2019. PMID: 31540982 Free PMC article.
-
Release Profiles of Carvacrol or Chlorhexidine of PLA/Graphene Nanoplatelets Membranes Prepared Using Electrospinning and Solution Blow Spinning: A Comparative Study.Molecules. 2023 Feb 19;28(4):1967. doi: 10.3390/molecules28041967. Molecules. 2023. PMID: 36838955 Free PMC article.
-
Aquatic Pseudomonads Inhibit Oomycete Plant Pathogens of Glycine max.Front Microbiol. 2018 May 29;9:1007. doi: 10.3389/fmicb.2018.01007. eCollection 2018. Front Microbiol. 2018. PMID: 29896163 Free PMC article.
References
-
- Zhao J, Schloss PD, Kalikin LM, Carmody LA, Foster BK, Petrosino JF, Cavalcoli JD, VanDevanter DR, Murray S, Li JZ, Young VB, LiPuma JJ. 2012. Decade-long bacterial community dynamics in cystic fibrosis airways. Proc Natl Acad Sci U S A 109:5809–5814. doi:10.1073/pnas.1120577109. - DOI - PMC - PubMed
-
- Jiricny N, Molin S, Foster K, Diggle SP, Scanlan PD, Ghoul M, Johansen HK, Santorelli LA, Popat R, West SA, Griffin AS. 2014. Loss of social behaviours in populations of Pseudomonas aeruginosa infecting lungs of patients with cystic fibrosis. PLoS One 9:e83124. doi:10.1371/journal.pone.0083124. - DOI - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
