

Reactive oxygen species drive evolution of pro-biofilm variants in pathogens by modulating cyclic-di-GMP levels
- PMID: 27881736
- PMCID: PMC5133437
- DOI: 10.1098/rsob.160162
Reactive oxygen species drive evolution of pro-biofilm variants in pathogens by modulating cyclic-di-GMP levels
Erratum in
-
Correction to 'Reactive oxygen species drive evolution of pro-biofilm variants in pathogens by modulating cyclic-di-GMP levels'.Open Biol. 2017 Sep;7(9):170197. doi: 10.1098/rsob.170197. Open Biol. 2017. PMID: 28954819 Free PMC article. No abstract available.
Abstract
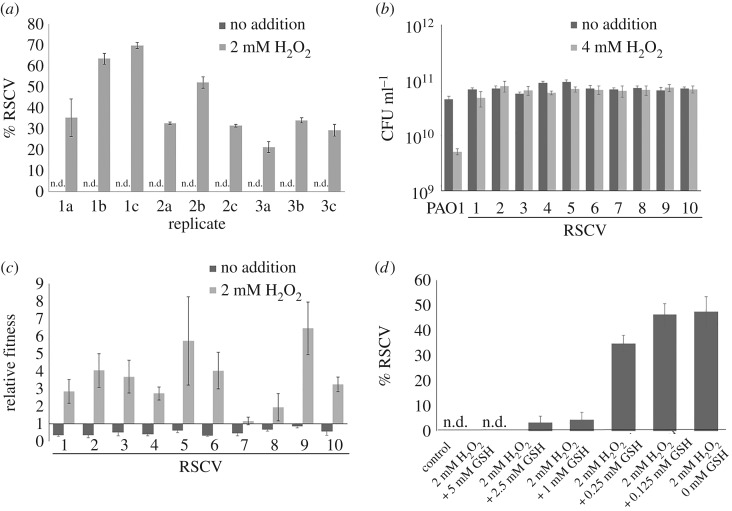
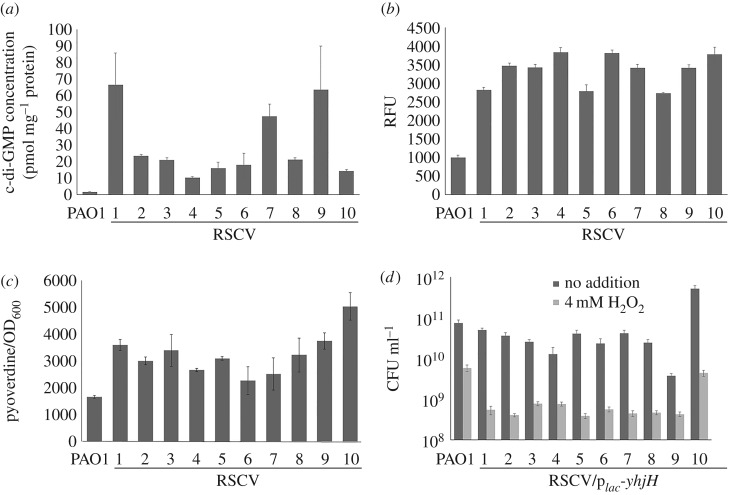
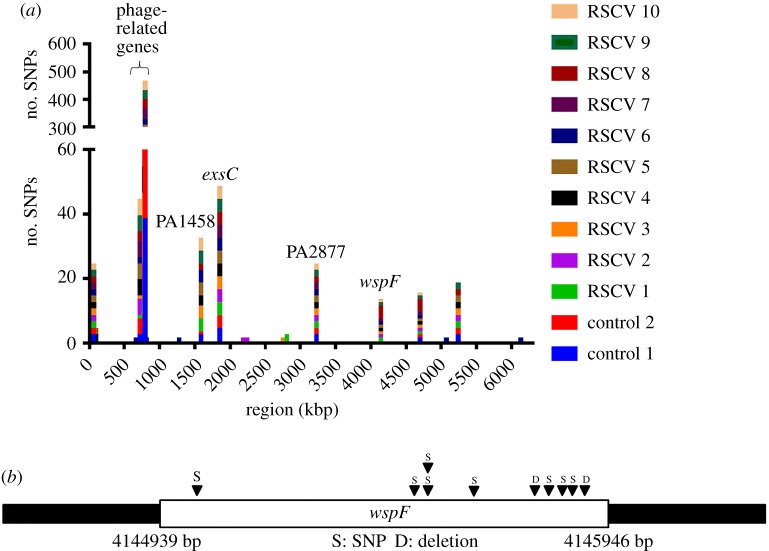
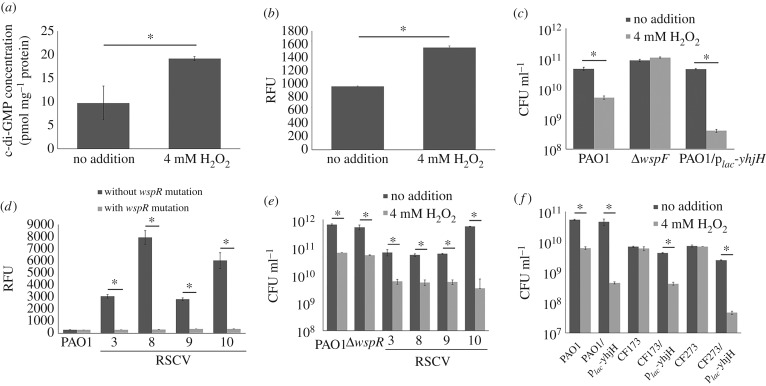
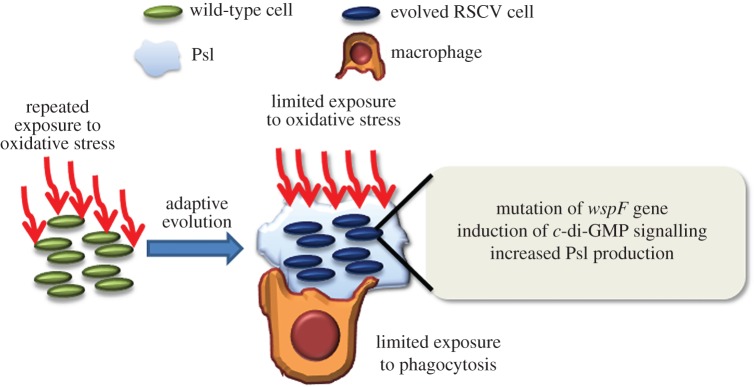
The host immune system offers a hostile environment with antimicrobials and reactive oxygen species (ROS) that are detrimental to bacterial pathogens, forcing them to adapt and evolve for survival. However, the contribution of oxidative stress to pathogen evolution remains elusive. Using an experimental evolution strategy, we show that exposure of the opportunistic pathogen Pseudomonas aeruginosa to sub-lethal hydrogen peroxide (H2O2) levels over 120 generations led to the emergence of pro-biofilm rough small colony variants (RSCVs), which could be abrogated by l-glutathione antioxidants. Comparative genomic analysis of the RSCVs revealed that mutations in the wspF gene, which encodes for a repressor of WspR diguanylate cyclase (DGC), were responsible for increased intracellular cyclic-di-GMP content and production of Psl exopolysaccharide. Psl provides the first line of defence against ROS and macrophages, ensuring the survival fitness of RSCVs over wild-type P. aeruginosa Our study demonstrated that ROS is an essential driving force for the selection of pro-biofilm forming pathogenic variants. Understanding the fundamental mechanism of these genotypic and phenotypic adaptations will improve treatment strategies for combating chronic infections.
Keywords: Pseudomonas aeruginosa; adaptive evolution; biofilms; c-di-GMP; reactive oxygen species; rough small colony variants.
© 2016 The Authors.
Figures


References
-
- Bjarnsholt T, Jensen PO, Fiandaca MJ, Pedersen J, Hansen CR, Andersen CB, Pressler T, Givskov M, Hoiby N. 2009. Pseudomonas aeruginosa biofilms in the respiratory tract of cystic fibrosis patients. Pediatr. Pulmonol. 44, 547–558. (doi:10.1002/ppul.21011) - DOI - PubMed
-
- Young D, Hussell T, Dougan G. 2002. Chronic bacterial infections: living with unwanted guests. Nat. Immunol. 3, 1026–1032. (doi:10.1038/ni1102-1026) - DOI - PubMed
-
- Domenech M, Ramos-Sevillano E, Garcia E, Moscoso M, Yuste J. 2013. Biofilm formation avoids complement immunity and phagocytosis of Streptococcus pneumoniae. Infect. Immun. 81, 2606–2615. (doi:10.1128/IAI.00491-13) - DOI - PMC - PubMed
-
- Alhede M, Bjarnsholt T, Givskov M, Alhede M. 2014. Pseudomonas aeruginosa biofilms: mechanisms of immune evasion. Adv. Appl. Microbiol. 86, 1–40. (doi:10.1016/B978-0-12-800262-9.00001-9) - DOI - PubMed
-
- Chiang WC, Nilsson M, Jensen PO, Hoiby N, Nielsen TE, Givskov M, Tolker-Nielsen T. 2013. Extracellular DNA shields against aminoglycosides in Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 57, 2352–2361. (doi:10.1128/AAC.00001-13) - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
