

Activation of Ventral Tegmental Area 5-HT2C Receptors Reduces Incentive Motivation
- PMID: 27882999
- PMCID: PMC5362069
- DOI: 10.1038/npp.2016.264
Activation of Ventral Tegmental Area 5-HT2C Receptors Reduces Incentive Motivation
Abstract
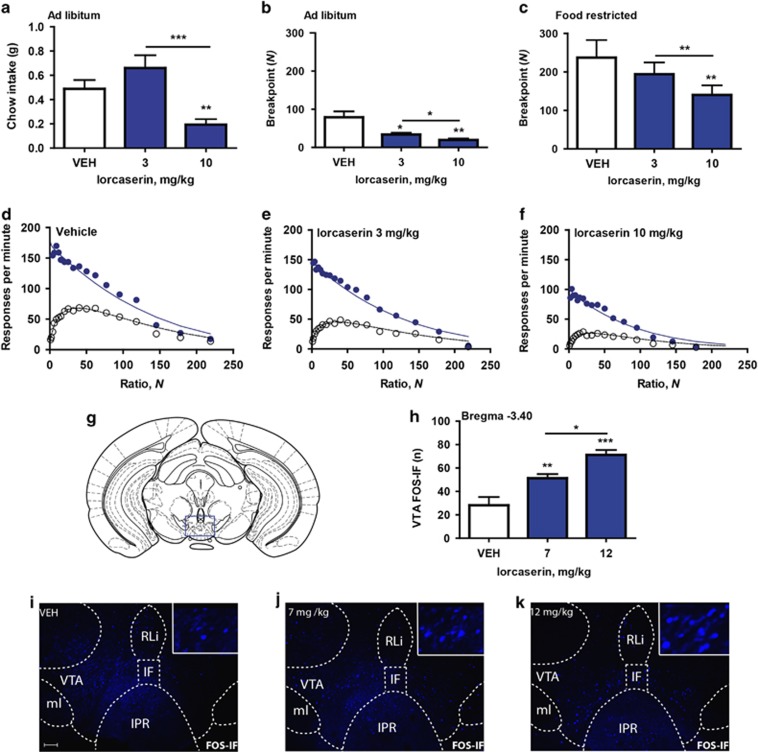
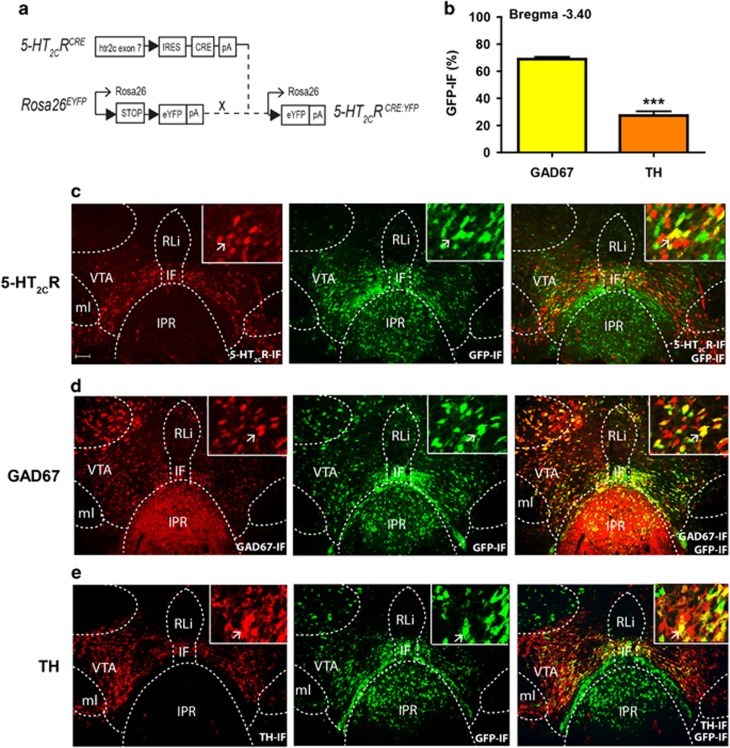
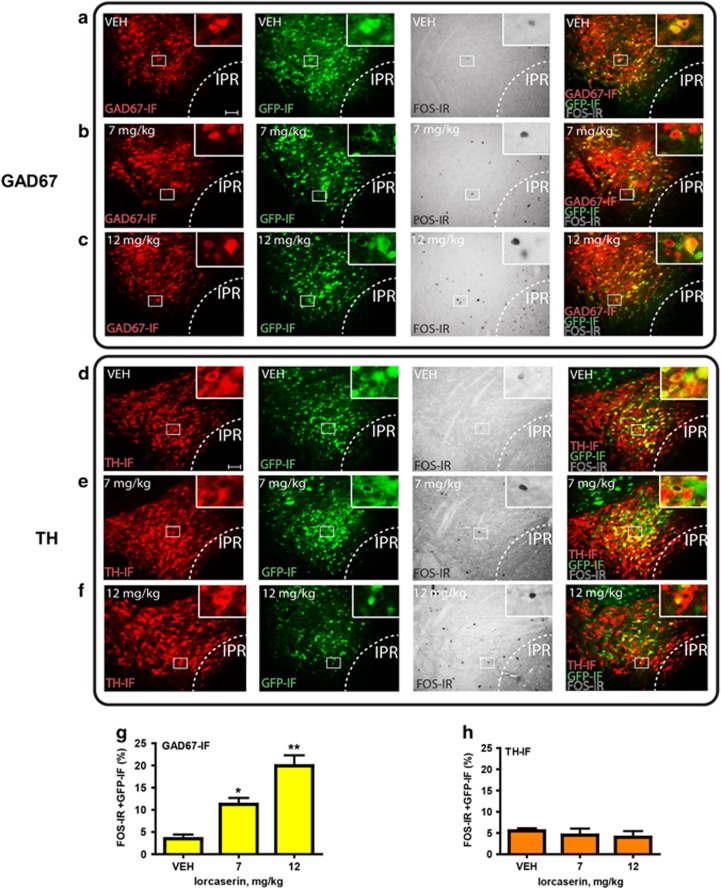
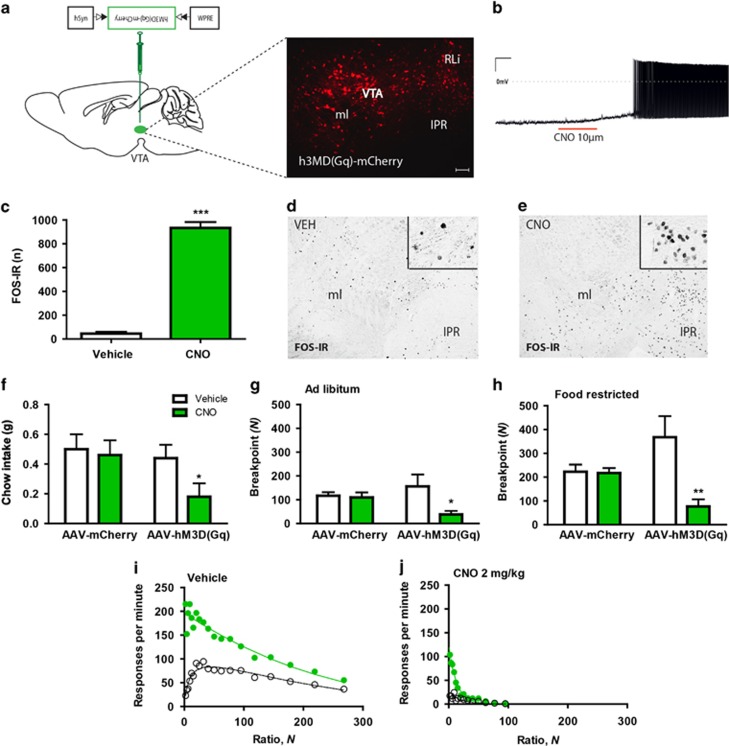
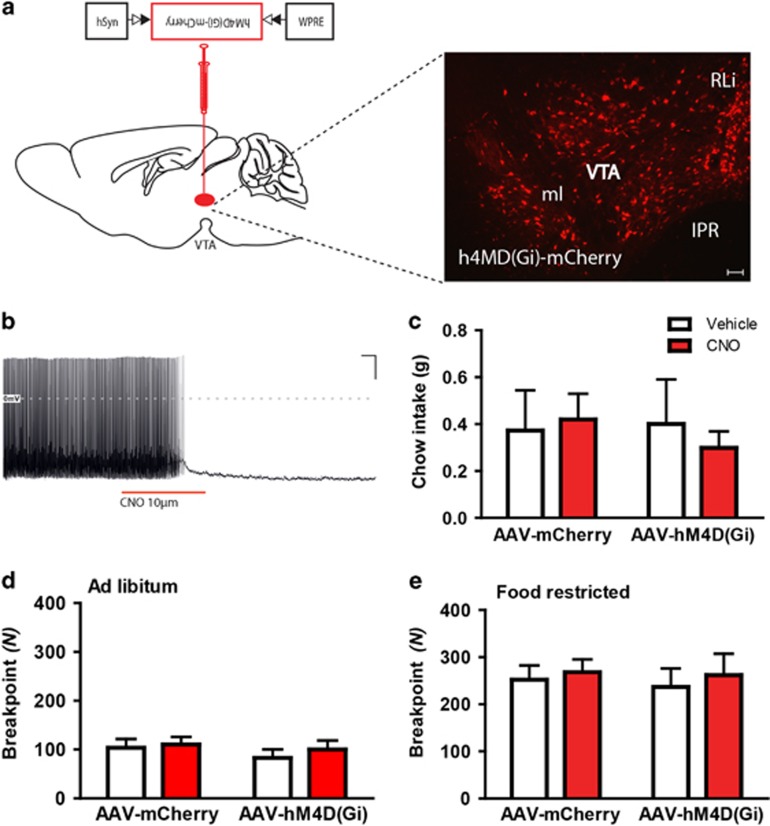
Obesity is primarily due to food intake in excess of the body's energetic requirements, intake that is not only associated with hunger but also the incentive value of food. The 5-hydroxytryptamine 2C receptor (5-HT2CR) is a target for the treatment of human obesity. Mechanistically, 5-HT2CRs are positioned to influence both homeostatic feeding circuits within the hypothalamus and reward circuits within the ventral tegmental area (VTA). Here we investigated the role of 5-HT2CRs in incentive motivation using a mathematical model of progressive ratio (PR) responding in mice. We found that the 5-HT2CR agonist lorcaserin significantly reduced both ad libitum chow intake and PR responding for chocolate pellets and increased c-fos expression in VTA 5-HT2CR expressing γ-aminobutyric acid (GABA) neurons, but not 5-HT2CR expressing dopamine (DA) neurons. We next adopted a chemogenetic approach using a 5-HT2CRCRE line to clarify the function of subset of 5-HT2C receptor expressing VTA neurons in the modulation of appetite and food-motivated behavior. Activation of VTA 5-HT2C receptor expressing neurons significantly reduced ad libitum chow intake, operant responding for chocolate pellets, and the incentive value of food. In contrast, chemogenetic inhibition of VTA 5-HT2C receptor expressing neurons had no effect on the feeding behavior. These results indicate that activation of the subpopulation of 5-HT2CR neurons within the VTA is sufficient to significantly reduce homeostatic feeding and effort-based intake of palatable food, and that this subset has an inhibitory role in motivational processes. These findings are relevant to the treatment of obesity.
Figures
References
-
- Aberman JE, Ward SJ, Salamone JD (1998). Effects of dopamine antagonists and accumbens dopamine depletions on time-constrained progressive-ratio performance. Pharmacol Biochem Behav 61: 341–348. - PubMed
-
- Bezzina G, Body S, Cheung TH, Hampson CL, Bradshaw CM, Glennon JC et al (2015). Evidence for a role of 5-HT2C receptors in the motor aspects of performance, but not the efficacy of food reinforcers, in a progressive ratio schedule. Psychopharmacology 232: 699–711. - PubMed
-
- Blundell JE (1986). Serotonin manipulations and the structure of feeding behaviour. Appetite 7 Suppl: 39–56. - PubMed
-
- Bradshaw CM, Killeen PR (2012). A theory of behaviour on progressive ratio schedules, with applications in behavioural pharmacology. Psychopharmacology 222: 549–564. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
