

A CRISPR-Cas9 Assisted Non-Homologous End-Joining Strategy for One-step Engineering of Bacterial Genome
- PMID: 27883076
- PMCID: PMC5121644
- DOI: 10.1038/srep37895
A CRISPR-Cas9 Assisted Non-Homologous End-Joining Strategy for One-step Engineering of Bacterial Genome
Abstract
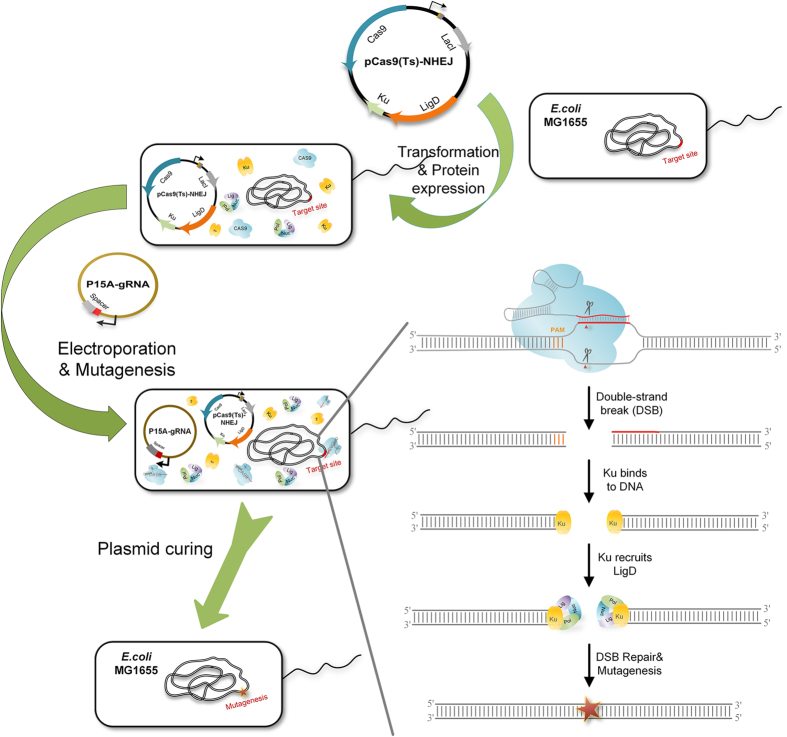
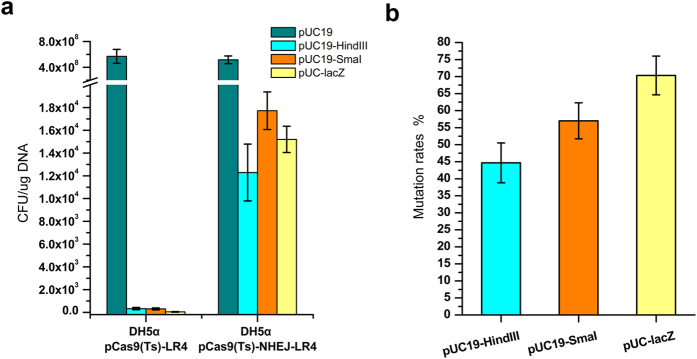
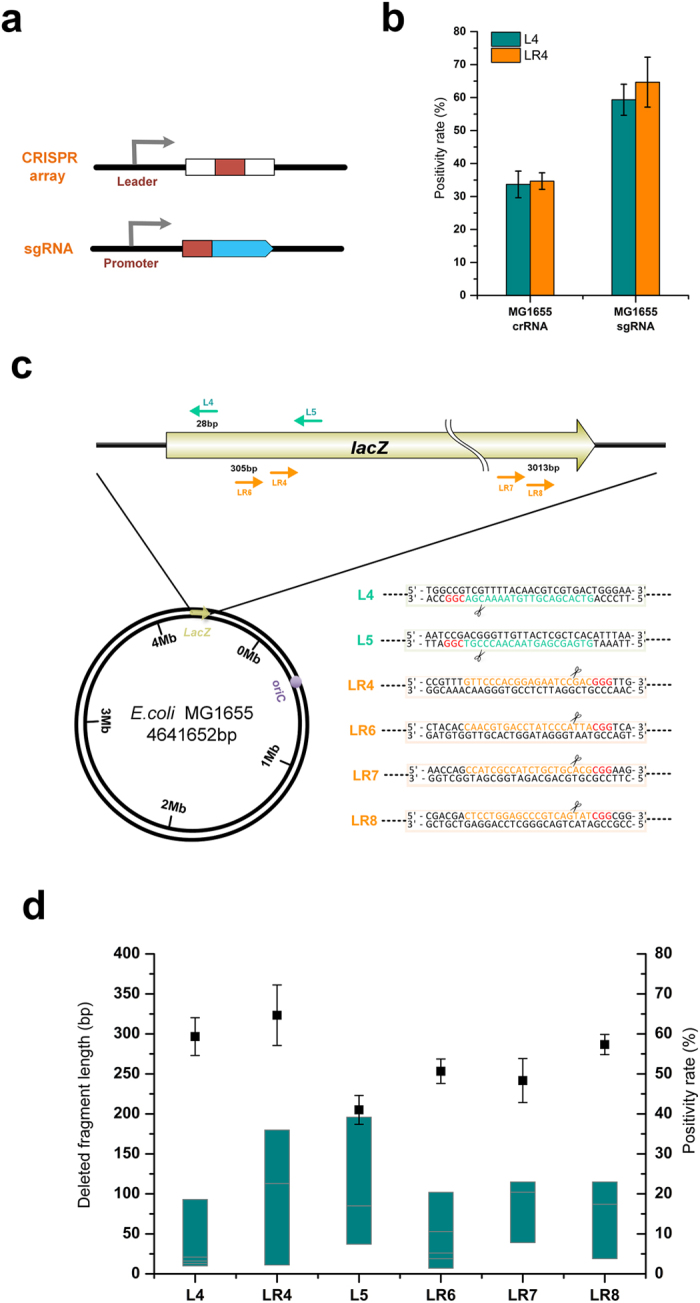
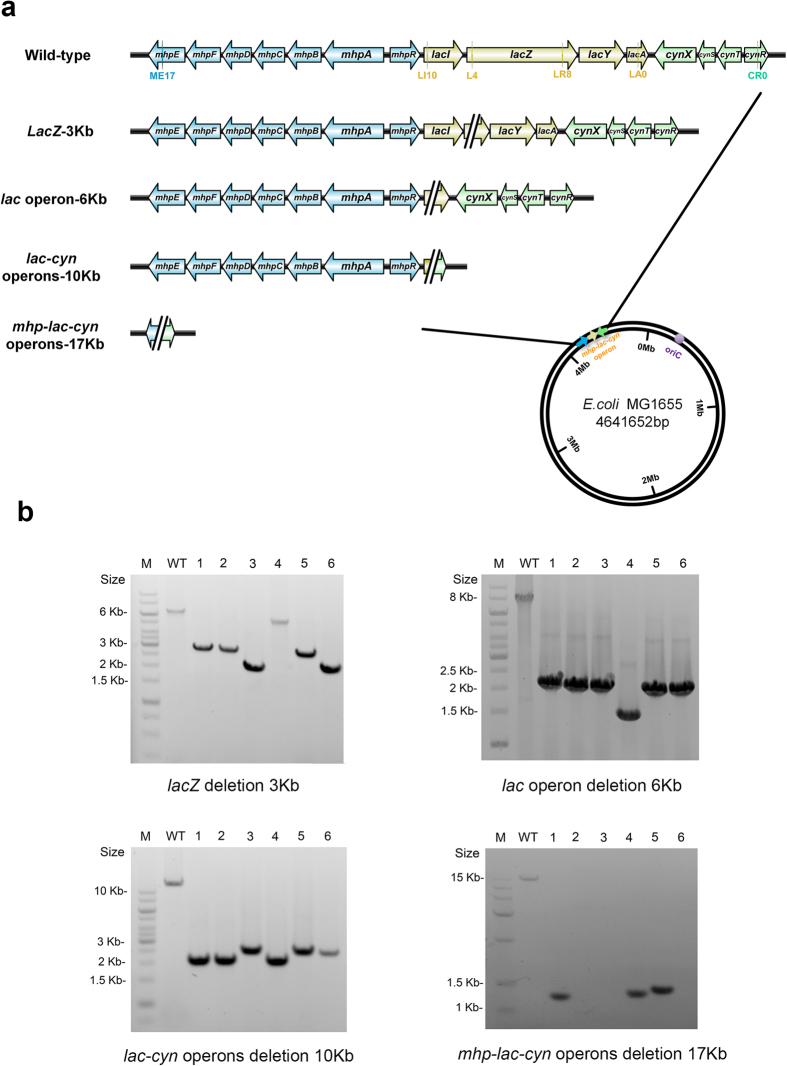
Homologous recombination-mediated genome engineering has been broadly applied in prokaryotes with high efficiency and accuracy. However, this method is limited in realizing larger-scale genome editing with numerous genes or large DNA fragments because of the relatively complicated procedure for DNA editing template construction. Here, we describe a CRISPR-Cas9 assisted non-homologous end-joining (CA-NHEJ) strategy for the rapid and efficient inactivation of bacterial gene (s) in a homologous recombination-independent manner and without the use of selective marker. Our study show that CA-NHEJ can be used to delete large chromosomal DNA fragments in a single step that does not require homologous DNA template. It is thus a novel and powerful tool for bacterial genomes reducing and possesses the potential for accelerating the genome evolution.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
