

Tapping the Brakes: Cellular and Synaptic Mechanisms that Regulate Thalamic Oscillations
- PMID: 27883901
- PMCID: PMC5131525
- DOI: 10.1016/j.neuron.2016.10.024
Tapping the Brakes: Cellular and Synaptic Mechanisms that Regulate Thalamic Oscillations
Abstract
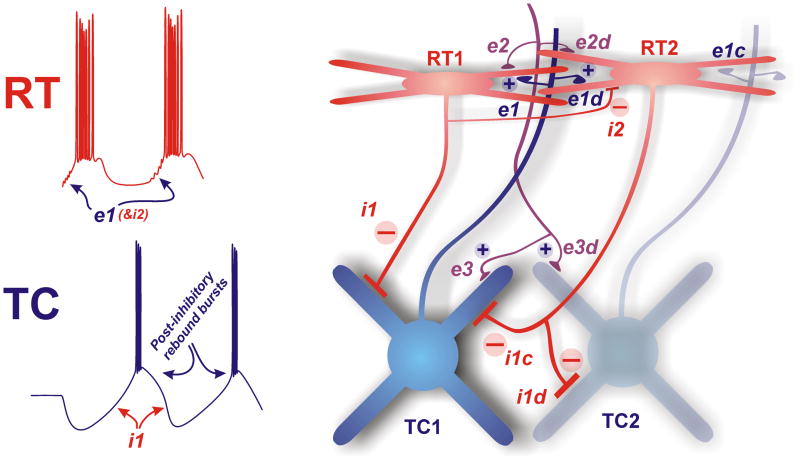
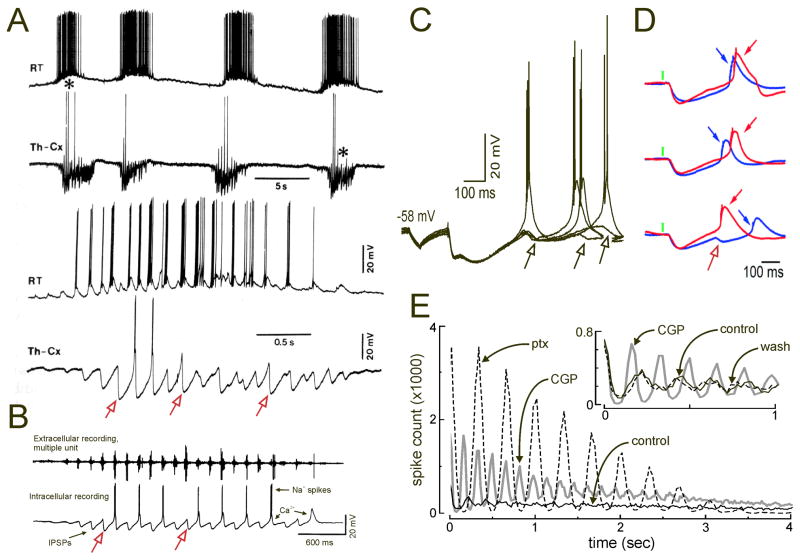
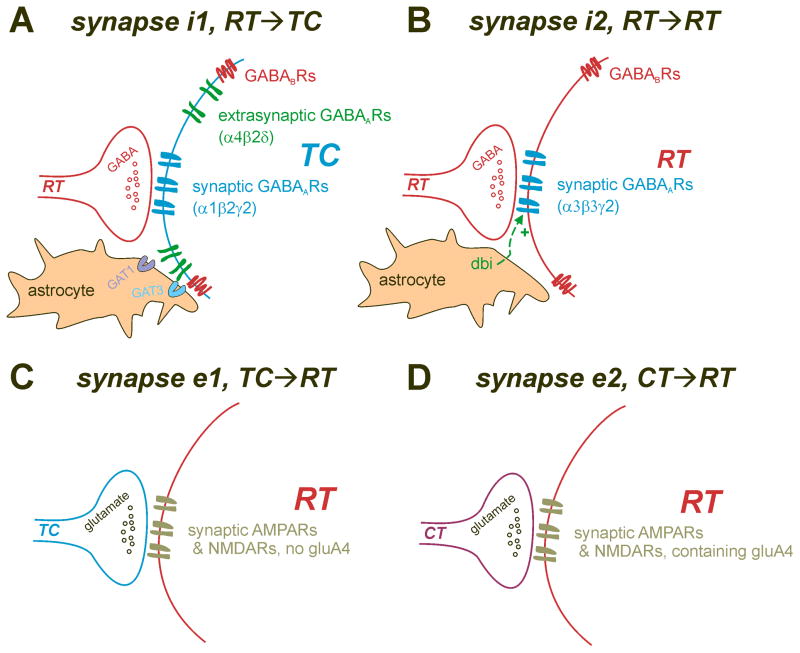
Thalamic oscillators contribute to both normal rhythms associated with sleep and anesthesia and abnormal, hypersynchronous oscillations that manifest behaviorally as absence seizures. In this review, we highlight new findings that refine thalamic contributions to cortical rhythms and suggest that thalamic oscillators may be subject to both local and global control. We describe endogenous thalamic mechanisms that limit network synchrony and discuss how these protective brakes might be restored to prevent absence seizures. Finally, we describe how intrinsic and circuit-level specializations among thalamocortical loops may determine their involvement in widespread oscillations and render subsets of thalamic nuclei especially vulnerable to pathological synchrony.
Keywords: absence epilepsy; oscillation; reticular nucleus; sparse networks; spindle; synchrony; thalamus.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Aker RG, Tezcan K, Çarçak N, Sakalli E, Akin D, Onat FY. Localized cortical injections of ethosuximide suppress spike-and-wave activity and reduce the resistance to kindling in genetic absence epilepsy rats (GAERS) Epilepsy Res. 2010;89:7–16. - PubMed
-
- Anderer P, Klösch G, Gruber G, Trenker E, Pascual-Marqui RD, Zeitlhofer J, Barbanoj MJ, Rappelsberger P, Saletu B. Low-resolution brain electromagnetic tomography revealed simultaneously active frontal and parietal sleep spindle sources in the human cortex. Neuroscience. 2001;103:581–592. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
