

Pathogens Inactivated by Low-Energy-Electron Irradiation Maintain Antigenic Properties and Induce Protective Immune Responses
- PMID: 27886076
- PMCID: PMC5127033
- DOI: 10.3390/v8110319
Pathogens Inactivated by Low-Energy-Electron Irradiation Maintain Antigenic Properties and Induce Protective Immune Responses
Abstract
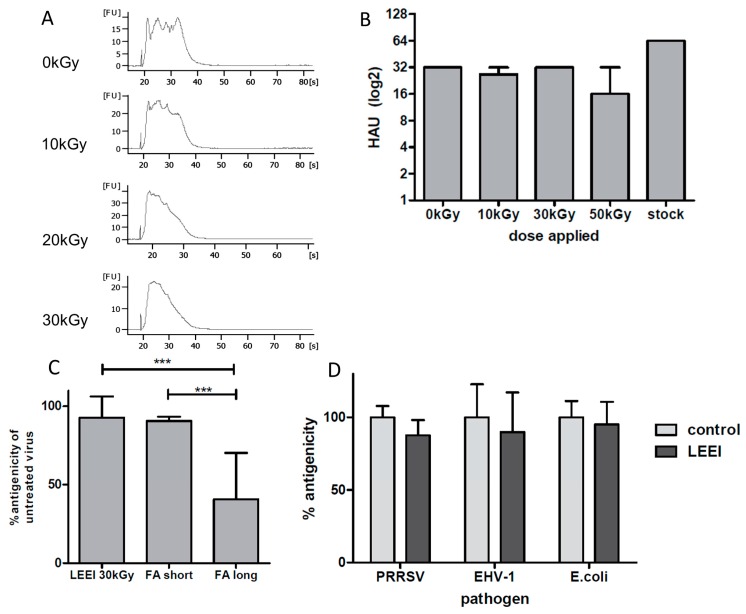
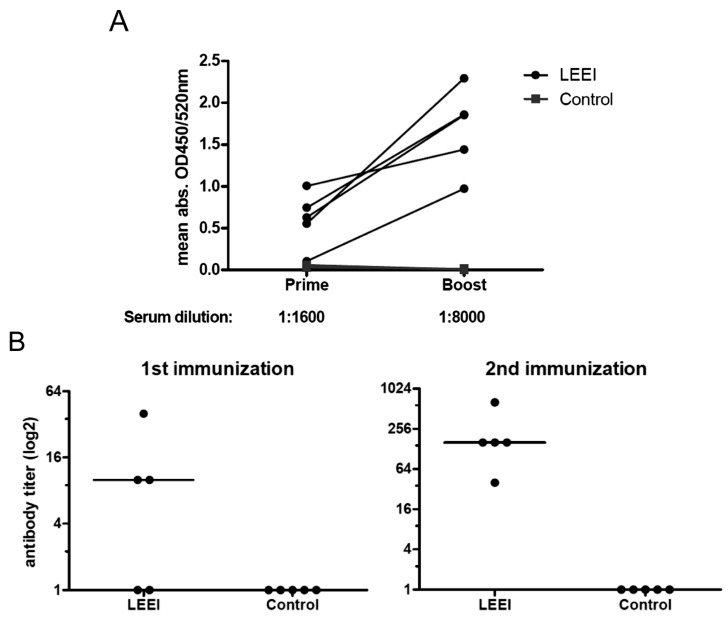
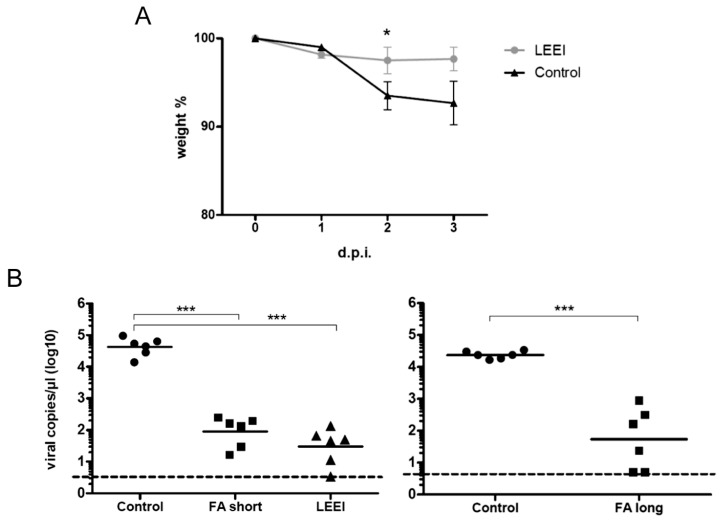
Inactivated vaccines are commonly produced by incubating pathogens with chemicals such as formaldehyde or β-propiolactone. This is a time-consuming process, the inactivation efficiency displays high variability and extensive downstream procedures are often required. Moreover, application of chemicals alters the antigenic components of the viruses or bacteria, resulting in reduced antibody specificity and therefore stimulation of a less effective immune response. An alternative method for inactivation of pathogens is ionizing radiation. It acts very fast and predominantly damages nucleic acids, conserving most of the antigenic structures. However, currently used irradiation technologies (mostly gamma-rays and high energy electrons) require large and complex shielding constructions to protect the environment from radioactivity or X-rays generated during the process. This excludes them from direct integration into biological production facilities. Here, low-energy electron irradiation (LEEI) is presented as an alternative inactivation method for pathogens in liquid solutions. LEEI can be used in normal laboratories, including good manufacturing practice (GMP)- or high biosafety level (BSL)-environments, as only minor shielding is necessary. We show that LEEI efficiently inactivates different viruses (influenza A (H3N8), porcine reproductive and respiratory syndrome virus (PRRSV), equine herpesvirus 1 (EHV-1)) and bacteria (Escherichia coli) and maintains their antigenicity. Moreover, LEEI-inactivated influenza A viruses elicit protective immune responses in animals, as analyzed by virus neutralization assays and viral load determination upon challenge. These results have implications for novel ways of developing and manufacturing inactivated vaccines with improved efficacy.
Keywords: influenza A; low-energy electron irradiation; pathogen inactivation; vaccines.
Conflict of interest statement
The technology described in this manuscript is part of an IP application (WO 2015011265).
Figures
Similar articles
-
Automated application of low energy electron irradiation enables inactivation of pathogen- and cell-containing liquids in biomedical research and production facilities.Sci Rep. 2020 Jul 30;10(1):12786. doi: 10.1038/s41598-020-69347-7. Sci Rep. 2020. PMID: 32732876 Free PMC article.
-
Low-Energy Electron Irradiation of Tick-Borne Encephalitis Virus Provides a Protective Inactivated Vaccine.Front Immunol. 2022 Mar 7;13:825702. doi: 10.3389/fimmu.2022.825702. eCollection 2022. Front Immunol. 2022. PMID: 35340807 Free PMC article.
-
Low-Energy Electron Irradiation (LEEI) for the Generation of Inactivated Bacterial Vaccines.Methods Mol Biol. 2022;2414:97-113. doi: 10.1007/978-1-0716-1900-1_7. Methods Mol Biol. 2022. PMID: 34784034
-
Inactivated and subunit vaccines against porcine reproductive and respiratory syndrome: Current status and future direction.Vaccine. 2015 Jun 17;33(27):3065-72. doi: 10.1016/j.vaccine.2015.04.102. Epub 2015 May 14. Vaccine. 2015. PMID: 25980425 Review.
-
Photochemical inactivation of viruses with psoralens: an overview.Blood Cells. 1992;18(1):7-25. Blood Cells. 1992. PMID: 1617194 Review.
Cited by
-
Viral inactivation by light.eLight. 2022;2(1):18. doi: 10.1186/s43593-022-00029-9. Epub 2022 Sep 26. eLight. 2022. PMID: 36187558 Free PMC article. Review.
-
Stabilization and Sterilization of Pericardial Scaffolds by Ultraviolet and Low-Energy Electron Irradiation.Tissue Eng Part C Methods. 2018 Dec;24(12):717-729. doi: 10.1089/ten.TEC.2018.0285. Tissue Eng Part C Methods. 2018. PMID: 30412035 Free PMC article.
-
Influenza Virus Inactivated by Heavy Ion Beam Irradiation Stimulates Antigen-Specific Immune Responses.Pharmaceutics. 2024 Mar 27;16(4):465. doi: 10.3390/pharmaceutics16040465. Pharmaceutics. 2024. PMID: 38675126 Free PMC article.
-
Electricity-producing Staphylococcus epidermidis counteracts Cutibacterium acnes.Sci Rep. 2021 Jun 7;11(1):12001. doi: 10.1038/s41598-021-91398-7. Sci Rep. 2021. PMID: 34099817 Free PMC article.
-
Low Energy Electron Irradiation Is a Potent Alternative to Gamma Irradiation for the Inactivation of (CAR-)NK-92 Cells in ATMP Manufacturing.Front Immunol. 2021 Jun 4;12:684052. doi: 10.3389/fimmu.2021.684052. eCollection 2021. Front Immunol. 2021. PMID: 34149724 Free PMC article.
References
-
- Gomez P.L., Robinson J.M., Rogalewicz J.A. Vaccine manufacturing. In: Plotkin S.A., Orenstein W.A., Offit P.A., editors. Vaccines. 6th ed. W.B. Saunders; London, UK: 2013. pp. 44–57.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
