

Nonreciprocal complementation of KNOX gene function in land plants
- PMID: 27886385
- PMCID: PMC5637896
- DOI: 10.1111/nph.14318
Nonreciprocal complementation of KNOX gene function in land plants
Abstract
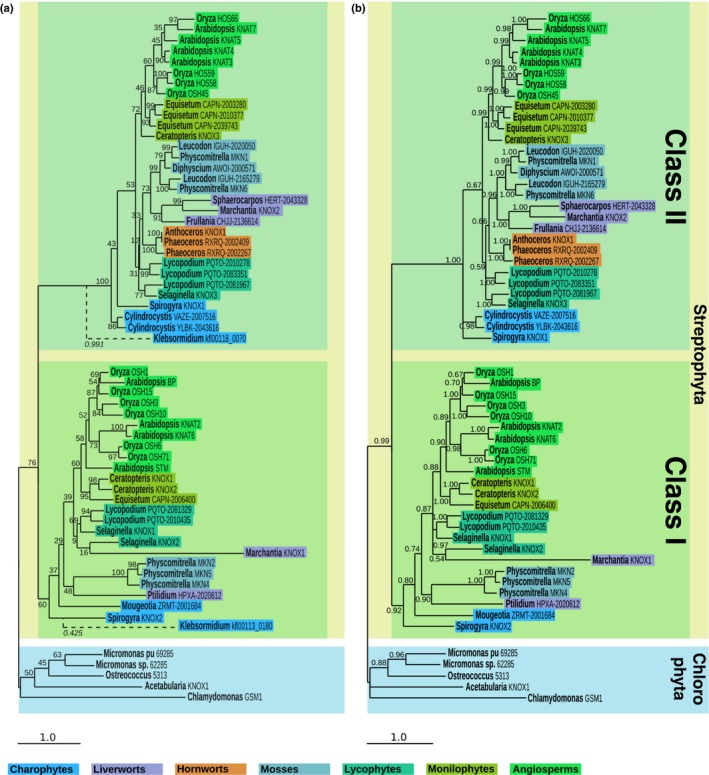
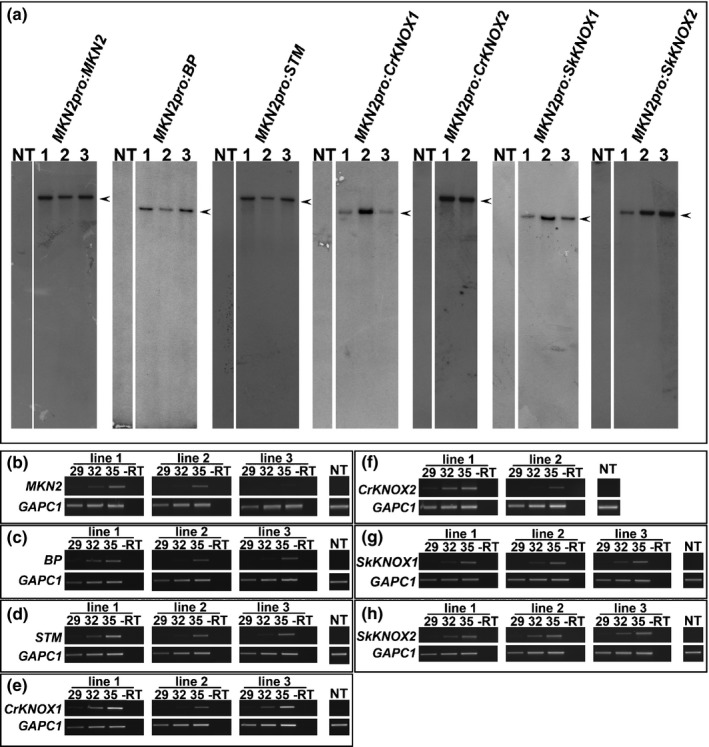
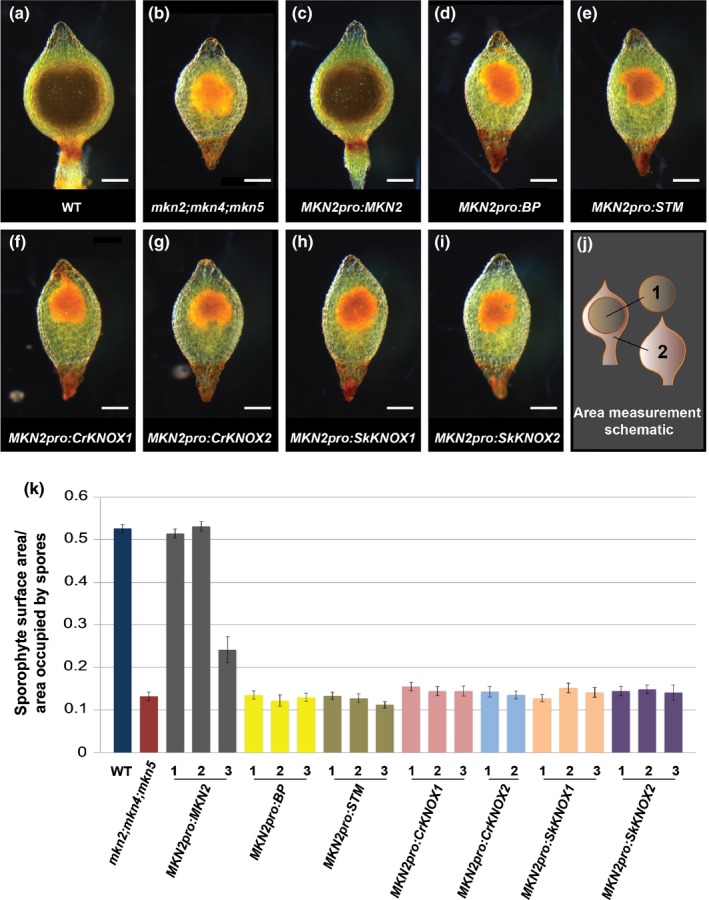
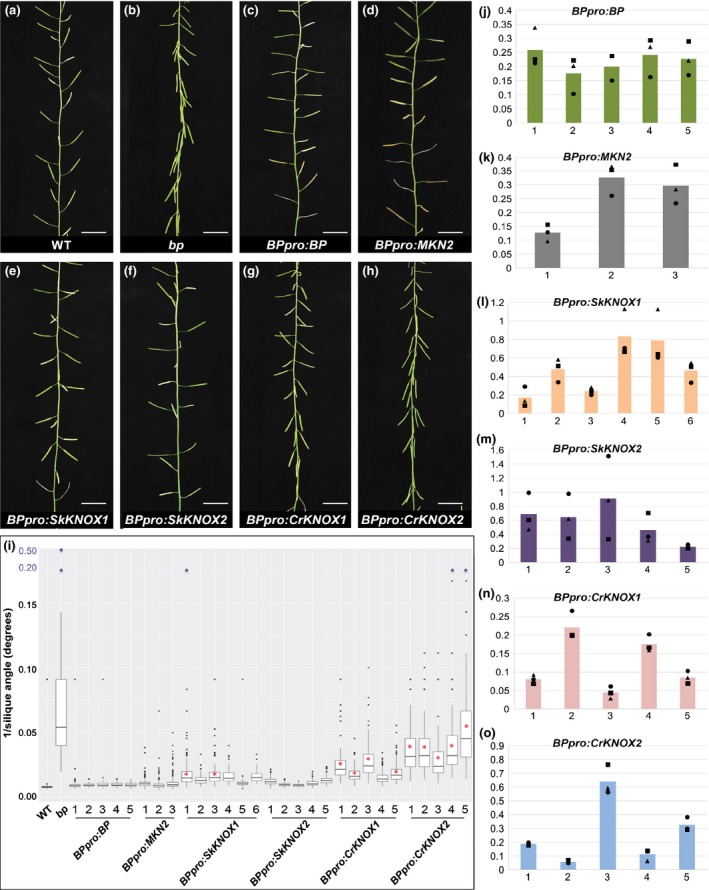
Class I KNOTTED-LIKE HOMEOBOX (KNOX) proteins regulate development of the multicellular diploid sporophyte in both mosses and flowering plants; however, the morphological context in which they function differs. In order to determine how Class I KNOX function was modified as land plants evolved, phylogenetic analyses and cross-species complementation assays were performed. Our data reveal that a duplication within the charophyte sister group to land plants led to distinct Class I and Class II KNOX gene families. Subsequently, Class I sequences diverged substantially in the nonvascular bryophyte groups (liverworts, mosses and hornworts), with moss sequences being most similar to those in vascular plants. Despite this similarity, moss mutants were not complemented by vascular plant KNOX genes. Conversely, the Arabidopsis brevipedicellus (bp-9) mutant was complemented by the PpMKN2 gene from the moss Physcomitrella patens. Lycophyte KNOX genes also complemented bp-9 whereas fern genes only partially complemented the mutant. This lycophyte/fern distinction is mirrored in the phylogeny of KNOX-interacting BELL proteins, in that a gene duplication occurred after divergence of the two groups. Together, our results imply that the moss MKN2 protein can function in a broader developmental context than vascular plant KNOX proteins, the narrower scope having evolved progressively as lycophytes, ferns and flowering plants diverged.
Keywords: Ceratopteris richardii; Physcomitrella patens; Selaginella kraussiana; Arabidopsis; KNOTTED homeobox genes; cross-species complementation; land plant evolution; phylogeny.
© 2016 The Authors. New Phytologist © 2016 New Phytologist Trust.
Figures
References
-
- Barton MK. 2010. Twenty years on: the inner workings of the shoot apical meristem, a developmental dynamo. Developmental Biology 341: 95–113. - PubMed
-
- Barton MK, Poethig RS. 1993. Formation of the shoot apical meristem in Arabidopsis thaliana: an analysis of development in the wild type and in the shoot meristemless mutant. Development 119: 823–831.
-
- Bravo‐Garcia A, Yasumura Y, Langdale JA. 2009. Specialization of the Golden2‐like regulatory pathway during land plant evolution. New Phytologist 183: 133–141. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
