

Drosophila Sulf1 is required for the termination of intestinal stem cell division during regeneration
- PMID: 27888216
- PMCID: PMC5278672
- DOI: 10.1242/jcs.195305
Drosophila Sulf1 is required for the termination of intestinal stem cell division during regeneration
Abstract
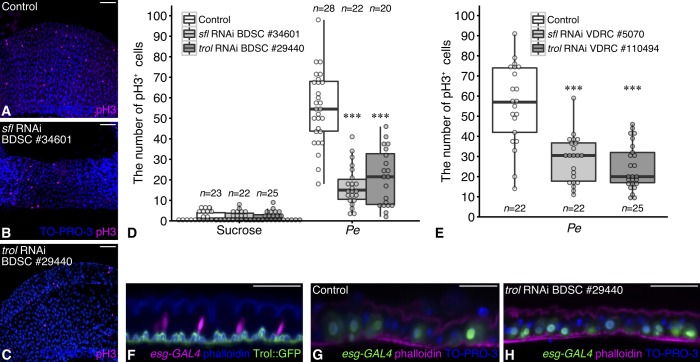
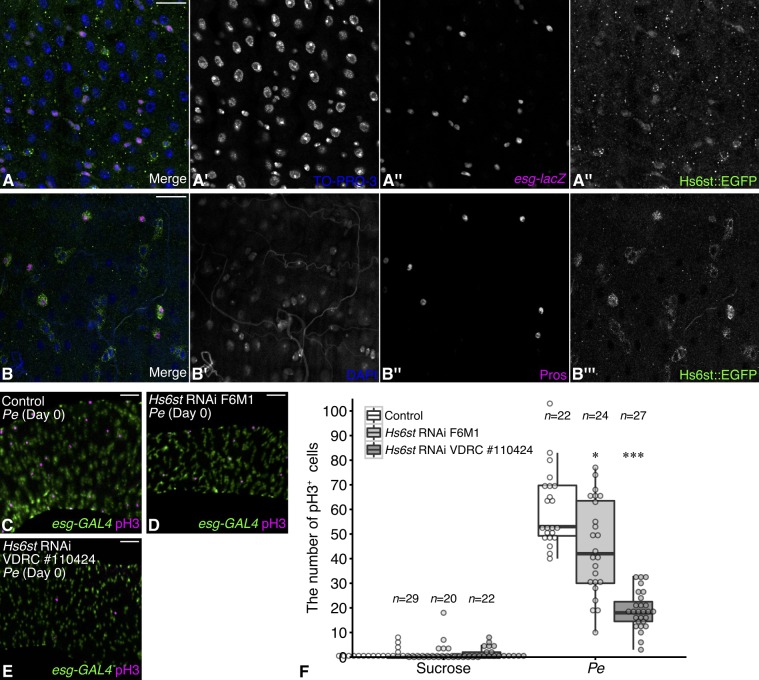
Stem cell division is activated to trigger regeneration in response to tissue damage. The molecular mechanisms by which this stem cell mitotic activity is properly repressed at the end of regeneration are poorly understood. Here, we show that a specific modification of heparan sulfate is crucial for regulating Drosophila intestinal stem cell (ISC) division during normal midgut homeostasis and regeneration. Loss of the extracellular heparan sulfate endosulfatase Sulf1 resulted in increased ISC division during normal homeostasis, which was caused by upregulation of mitogenic signaling including the JAK-STAT, EGFR and Hedgehog pathways. Using a regeneration model, we found that ISCs failed to properly halt division at the termination stage in Sulf1 mutants, showing that Sulf1 is required for terminating ISC division at the end of regeneration. We propose that post-transcriptional regulation of mitogen signaling by heparan sulfate structural modifications provides a new regulatory step for precise temporal control of stem cell activity during regeneration.
Keywords: Drosophila; Heparan sulfate proteoglycan; Intestine; Regeneration.
© 2017. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures






Similar articles
-
EGFR, Wingless and JAK/STAT signaling cooperatively maintain Drosophila intestinal stem cells.Dev Biol. 2011 Jun 1;354(1):31-43. doi: 10.1016/j.ydbio.2011.03.018. Epub 2011 Apr 1. Dev Biol. 2011. PMID: 21440535
-
The UPD3 cytokine couples environmental challenge and intestinal stem cell division through modulation of JAK/STAT signaling in the stem cell microenvironment.Dev Biol. 2013 Jan 15;373(2):383-93. doi: 10.1016/j.ydbio.2012.10.023. Epub 2012 Oct 27. Dev Biol. 2013. PMID: 23110761 Free PMC article.
-
Overexpression of dJmj differentially affects intestinal stem cells and differentiated enterocytes.Cell Signal. 2018 Jan;42:194-210. doi: 10.1016/j.cellsig.2017.10.017. Epub 2017 Nov 2. Cell Signal. 2018. PMID: 29102770
-
Intestinal stem cell response to injury: lessons from Drosophila.Cell Mol Life Sci. 2016 Sep;73(17):3337-49. doi: 10.1007/s00018-016-2235-9. Epub 2016 May 2. Cell Mol Life Sci. 2016. PMID: 27137186 Free PMC article. Review.
-
Intestinal stem cell asymmetric division in the Drosophila posterior midgut.J Cell Physiol. 2010 Sep;224(3):581-4. doi: 10.1002/jcp.22194. J Cell Physiol. 2010. PMID: 20578235 Review.
Cited by
-
Regulation of morphogen pathways by a Drosophila chondroitin sulfate proteoglycan Windpipe.J Cell Sci. 2023 Apr 1;136(7):jcs260525. doi: 10.1242/jcs.260525. Epub 2023 Apr 11. J Cell Sci. 2023. PMID: 36897575 Free PMC article.
-
Integration of Transcriptional Repression and Polycomb-Mediated Silencing of WUSCHEL in Floral Meristems.Plant Cell. 2019 Jul;31(7):1488-1505. doi: 10.1105/tpc.18.00450. Epub 2019 May 8. Plant Cell. 2019. PMID: 31068455 Free PMC article.
-
Heparan sulfate negatively regulates intestinal stem cell proliferation in Drosophila adult midgut.Biol Open. 2019 Oct 18;8(10):bio047126. doi: 10.1242/bio.047126. Biol Open. 2019. PMID: 31628141 Free PMC article.
-
Chondroitin sulfate proteoglycan Windpipe modulates Hedgehog signaling in Drosophila.Mol Biol Cell. 2020 Apr 1;31(8):813-824. doi: 10.1091/mbc.E19-06-0327. Epub 2020 Feb 12. Mol Biol Cell. 2020. PMID: 32049582 Free PMC article.
-
Refining the genomic profiles of North African sheep breeds through meta-analysis of worldwide genomic SNP data.Front Vet Sci. 2024 Feb 29;11:1339321. doi: 10.3389/fvets.2024.1339321. eCollection 2024. Front Vet Sci. 2024. PMID: 38487707 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
