

Amplification of USP13 drives ovarian cancer metabolism
- PMID: 27892457
- PMCID: PMC5133706
- DOI: 10.1038/ncomms13525
Amplification of USP13 drives ovarian cancer metabolism
Abstract
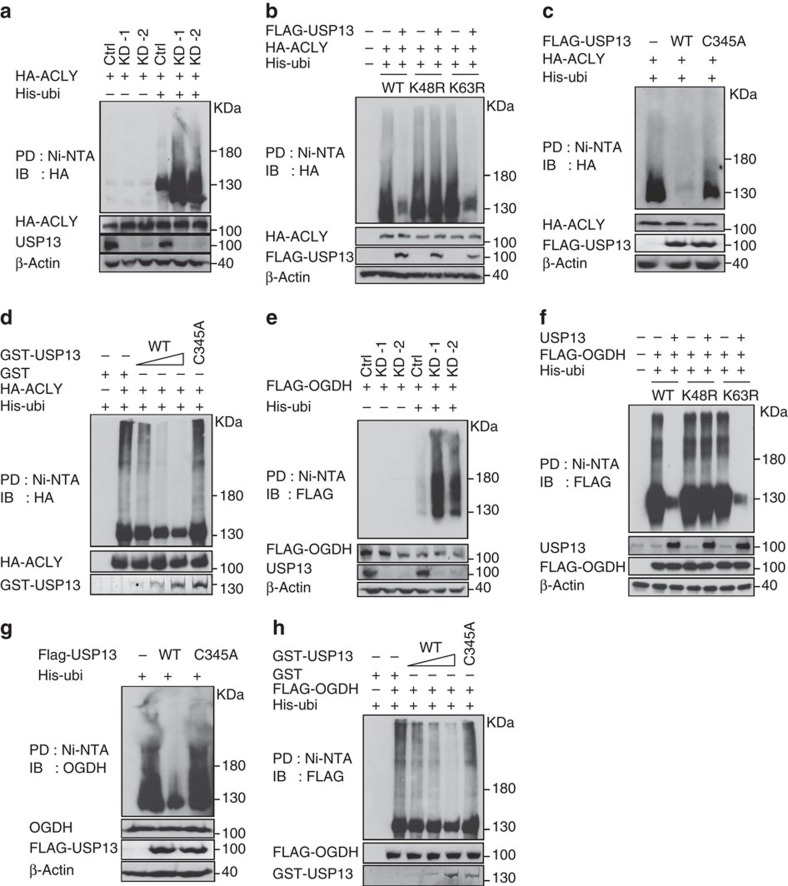
Dysregulated energetic metabolism has been recently identified as a hallmark of cancer. Although mutations in metabolic enzymes hardwire metabolism to tumourigenesis, they are relatively infrequent in ovarian cancer. More often, cancer metabolism is re-engineered by altered abundance and activity of the metabolic enzymes. Here we identify ubiquitin-specific peptidase 13 (USP13) as a master regulator that drives ovarian cancer metabolism. USP13 specifically deubiquitinates and thus upregulates ATP citrate lyase and oxoglutarate dehydrogenase, two key enzymes that determine mitochondrial respiration, glutaminolysis and fatty acid synthesis. The USP13 gene is co-amplified with PIK3CA in 29.3% of high-grade serous ovarian cancers and its overexpression is significantly associated with poor clinical outcome. Inhibiting USP13 remarkably suppresses ovarian tumour progression and sensitizes tumour cells to the treatment of PI3K/AKT inhibitor. Our results reveal an important metabolism-centric role of USP13, which may lead to potential therapeutics targeting USP13 in ovarian cancers.
Figures








Similar articles
-
USP13 promotes development and metastasis of high-grade serous ovarian carcinoma in a novel mouse model.Oncogene. 2022 Mar;41(13):1974-1985. doi: 10.1038/s41388-022-02224-x. Epub 2022 Feb 16. Oncogene. 2022. PMID: 35173307 Free PMC article.
-
Deubiquitylation and stabilization of PTEN by USP13.Nat Cell Biol. 2013 Dec;15(12):1486-1494. doi: 10.1038/ncb2874. Epub 2013 Nov 24. Nat Cell Biol. 2013. PMID: 24270891 Free PMC article.
-
USP13 regulates the RAP80-BRCA1 complex dependent DNA damage response.Nat Commun. 2017 Jun 1;8:15752. doi: 10.1038/ncomms15752. Nat Commun. 2017. PMID: 28569838 Free PMC article.
-
USP13: A therapeutic target for combating tumorigenesis and antitumor therapy resistance.Int J Biol Macromol. 2025 Apr;304(Pt 1):140608. doi: 10.1016/j.ijbiomac.2025.140608. Epub 2025 Feb 1. Int J Biol Macromol. 2025. PMID: 39900156 Review.
-
PI3K/AKT/mTOR signaling pathway as a therapeutic target for ovarian cancer.Arch Gynecol Obstet. 2014 Dec;290(6):1067-78. doi: 10.1007/s00404-014-3377-3. Epub 2014 Aug 3. Arch Gynecol Obstet. 2014. PMID: 25086744 Review.
Cited by
-
Mutational Landscape of Esophageal Squamous Cell Carcinoma in an Indian Cohort.Front Oncol. 2020 Aug 20;10:1457. doi: 10.3389/fonc.2020.01457. eCollection 2020. Front Oncol. 2020. PMID: 32974170 Free PMC article.
-
USP13 promotes development and metastasis of high-grade serous ovarian carcinoma in a novel mouse model.Oncogene. 2022 Mar;41(13):1974-1985. doi: 10.1038/s41388-022-02224-x. Epub 2022 Feb 16. Oncogene. 2022. PMID: 35173307 Free PMC article.
-
N6-methyladenosine-modified USP13 induces pro-survival autophagy and imatinib resistance via regulating the stabilization of autophagy-related protein 5 in gastrointestinal stromal tumors.Cell Death Differ. 2023 Feb;30(2):544-559. doi: 10.1038/s41418-022-01107-8. Epub 2022 Dec 17. Cell Death Differ. 2023. PMID: 36528756 Free PMC article.
-
The emerging role of deubiquitylating enzymes as therapeutic targets in cancer metabolism.Cancer Cell Int. 2022 Mar 20;22(1):130. doi: 10.1186/s12935-022-02524-y. Cancer Cell Int. 2022. PMID: 35307036 Free PMC article. Review.
-
USP13 serves as a tumor suppressor via the PTEN/AKT pathway in oral squamous cell carcinoma.Cancer Manag Res. 2019 Oct 31;11:9175-9183. doi: 10.2147/CMAR.S186829. eCollection 2019. Cancer Manag Res. 2019. PMID: 31802942 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
